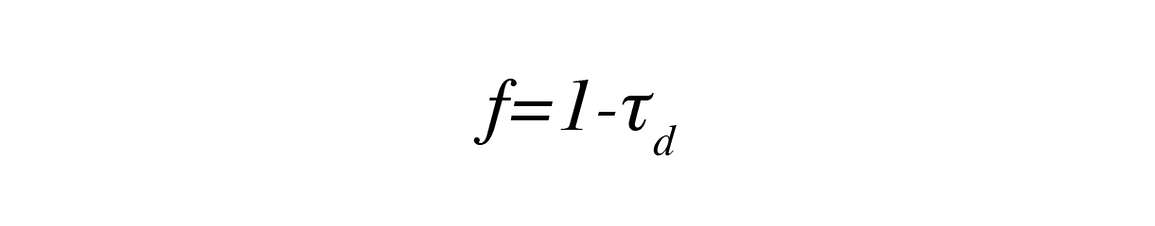Zapoznaj się z artykułami, podcastami, studiami przypadków i najlepszymi praktykami napisanymi przez naukowców, którzy spędzili dziesięciolecia w terenie. Od fizyki gleby po konfigurację czujników, jest to źródło praktycznych wskazówek badawczych.
Uzyskaj jasne, bezpośrednie odpowiedzi od zespołu, który rozumie Twoje metody i misję. Niezależnie od tego, czy potrzebujesz filmów instruktażowych, wskazówek dotyczących rozwiązywania problemów czy wiedzy technicznej, jesteśmy tutaj, aby uprościć złożoność.
Od kalibracji czujników po laboratoryjne usługi pomiarowe, nasz zespół pomaga zbierać istotne dane z precyzją i wydajnością - dzięki czemu możesz skupić się na nauce, a nie na konfiguracji.
Wypełnij poniższy formularz, aby pomóc nam połączyć Cię z odpowiednim ekspertem. Przygotujemy wymagane informacje, a następnie skontaktujemy się z Tobą tak szybko, jak to możliwe.
Kompletny przewodnik badacza po Leaf Area Index (LAI)
Leaf area index to pojedyncza liczba
-statystyczna migawka okapu wykonana w określonym czasie. Ale ta jedna liczba może prowadzić do istotnych wniosków.
Leaf area index (LAI) jest jednym z najczęściej stosowanych pomiarów opisujących strukturę korony roślin. LAI jest również przydatny do zrozumienia funkcji korony, ponieważ wiele wymian masy i energii między biosferą a atmosferą zachodzi na powierzchni liści. Z tych powodów LAI jest często kluczową zmienną biofizyczną wykorzystywaną w modelach biogeochemicznych, hydrologicznych i ekologicznych. Leaf area index jest również powszechnie stosowany jako miara wzrostu i produktywności upraw i lasów w skali przestrzennej od działki do globu. W tym artykule dowiesz się, jak zmierzyć leaf area index, co to jest i jak go używać.
W przeszłości pomiar leaf area index (LAI) był trudny i czasochłonny. Jednak teoria i technologia opracowane w ostatnich latach sprawiły, że pomiar LAI jest znacznie prostszy i bardziej wykonalny dla szerokiej gamy zadaszeń. Pobierz ten przewodnik po zastosowaniach, aby uzyskać krótkie wprowadzenie do teorii i instrumentów używanych do pomiaru leaf area index. Omówiono kilka scenariuszy i specjalnych rozważań, które pomogą osobom wybrać i zastosować najbardziej odpowiednią metodę dla ich potrzeb badawczych.
Co to jest leaf area index (LAI)?
Leaf area index (LAI) określa ilość materiału liściowego w łanie. Z definicji jest to stosunek jednostronnej powierzchni liści na jednostkę powierzchni gruntu. LAI jest bezjednostkowy, ponieważ jest to stosunek powierzchni. Na przykład łan o LAI równym 1 ma stosunek powierzchni liści do powierzchni gruntu 1:1 (rysunek 1a). Korona o leaf area index równym 3 miałaby stosunek powierzchni liści do powierzchni gruntu wynoszący 3:1 (rysunek 1b).
W skali globalnej LAI jest bardzo zróżnicowany. leaf area index Niektóre ekosystemy pustynne mają LAI mniejszy niż 1, podczas gdy najgęstsze lasy tropikalne mogą mieć LAI nawet 9. Lasy i krzewy na średnich szerokościach geograficznych mają zwykle wartości LAI między 3 a 6.
Sezonowo, jednoroczne i liściaste zadaszenia i pola uprawne mogą wykazywać duże wahania LAI. Na przykład, od siewu do dojrzałości, kukurydza leaf area index może wahać się od 0 do 6. Oczywiście LAI jest użyteczną miarą do opisywania zarówno przestrzennych, jak i czasowych wzorców wzrostu i produktywności koron drzew.
Dowiedz się więcej o podstawach leaf area index (LAI) w poniższym filmie. Dr Steve Garrity omawia teorię stojącą za pomiarem, metody bezpośrednie i pośrednie, zmienność między tymi metodami, rzeczy, które należy wziąć pod uwagę przy wyborze metody oraz zastosowania leaf area index liści.
Jak mierzyć leaf area index
Nie ma jednego najlepszego sposobu pomiaru LAI. Każda metoda ma zalety i wady. Wybrana metoda będzie w dużej mierze zależeć od celów badawczych. Badacz, który potrzebuje pojedynczego oszacowania LAI, może użyć innej metody niż ten, który monitoruje zmiany w leaf area index w czasie. Na przykład badacz użytków zielonych może preferować inną metodę niż badacz leśnictwa. W tym przewodniku omówimy podstawy teoretyczne każdej z głównych metod wraz z kluczowymi zaletami i ograniczeniami.
Pomiar bezpośredni
Tradycyjnie badacze mierzyli leaf area index zbierając wszystkie liście z poletka i skrupulatnie mierząc powierzchnię każdego liścia. Nowoczesny sprzęt, taki jak skanery płaskie, sprawił, że proces ten stał się bardziej wydajny, ale nadal jest pracochłonny, czasochłonny i destrukcyjny. W wysokich koronach drzew może to być nawet niewykonalne. Pozostaje to jednak najdokładniejszą metodą obliczania leaf area index , ponieważ każdy pojedynczy liść jest fizycznie mierzony. Pułapki na ściółkę są kolejnym sposobem bezpośredniego pomiaru LAI, ale nie działają dobrze w wiecznie zielonych koronach drzew i mogą przechwytywać informacje tylko z liści, które uschły i oderwały się od rośliny.
Pomiar pośredni
Kilkadziesiąt lat temu badacze koron drzew zaczęli szukać nowych sposobów pomiaru LAI, zarówno w celu zaoszczędzenia czasu, jak i uniknięcia niszczenia ekosystemów, które próbowali zmierzyć. Te pośrednie metody wywnioskowały LAI z pomiarów powiązanych zmiennych, takich jak ilość światła przepuszczanego lub odbijanego przez koronę.
Fotografia półkulista
Fotografia półkulista była jedną z pierwszych metod wykorzystywanych do pośredniego szacowania leaf area index. Badacze fotografowali baldachim z ziemi za pomocą obiektywu typu rybie oko. Fotografie były pierwotnie analizowane przez samych badaczy. Obecnie większość badaczy korzysta ze specjalistycznego oprogramowania do analizy obrazów i rozróżniania pikseli roślinnych i nieroślinnych.
Rysunek 2. Fotografia półsferyczna z mieszanego lasu liściastego przy użyciu obiektywu typu rybie oko w aparacie cyfrowym
Zalety: Fotografia półsferyczna ma zdecydowane zalety. Po pierwsze, zapewnia więcej niż tylko pomiary leaf area index . Może również zapewnić pomiary czaszy, takie jak frakcja luki, czas i czas trwania prześwietlenia słonecznego oraz inne wskaźniki architektury czaszy. Po drugie, obrazy baldachimu mogą być archiwizowane do późniejszego wykorzystania lub do ponownej analizy w miarę zmiany metod i ulepszania oprogramowania.
Ograniczenia: Fotografia półsferyczna ma jednak wady. Pomimo faktu, że obrazy są obecnie przetwarzane cyfrowo, subiektywność użytkownika pozostaje istotną kwestią. Użytkownicy muszą wybrać progi jasności obrazu, które odróżniają piksele nieba od pikseli roślinności, co powoduje, że wartości LAI różnią się w zależności od użytkownika lub przy użyciu różnych algorytmów analizy obrazu.
Fotografia półsferyczna jest również czasochłonna. Uzyskanie dobrej jakości zdjęć w terenie wymaga czasu, a ich analiza w laboratorium zajmuje jeszcze więcej czasu. Ponadto podczas wykonywania zdjęć niebo musi być jednolicie zachmurzone. Fotografia półsferyczna nie sprawdza się dobrze w przypadku krótkich łanów, takich jak pszenica i kukurydza, ponieważ korpus aparatu, obiektyw i statyw mogą fizycznie nie zmieścić się pod łanem.
Uwaga: Dla niektórych użytkowników instrumenty mierzące PAR oferują skrót. Niektóre modele wykorzystują wartości LAI do szacowania PAR. W takim przypadku przyrząd PAR może być użyty do bezpośredniego oszacowania poziomów PAR poniżej korony, poprawiając dokładność modelu.
Przepuszczalność promieniowania
Kilka dostępnych na rynku przyrządów, w tym METER LP-80 stanowią alternatywę dla fotografii hemisferycznej. Szacują one LAI na podstawie ilości energii świetlnej przepuszczanej przez korony roślin. Pomysł jest dość prosty; bardzo gęsty baldachim pochłonie więcej światła niż rzadki baldachim. Oznacza to, że musi istnieć jakiś związek między LAI a przechwytywaniem światła. Prawo Beera stanowi teoretyczną podstawę tego związku. Dla celów biofizyki środowiskowej prawo Beera jest sformułowane jako
gdzie PARt to transmitowane promieniowanie fotosyntetycznie czynne (PAR) mierzone w pobliżu powierzchni gruntu, PARi to PAR padające na wierzchołek czaszy, z to długość drogi fotonów przez ośrodek tłumiący, a k to współczynnik ekstynkcji. W przypadku baldachimów roślinnych, z odpowiada LAI, ponieważ liście są medium, przez które fotony są tłumione. Można zauważyć, że jeśli znamy k i mierzymy PARt i PARi, może być możliwe odwrócenie równania 1 w celu obliczenia z jako oszacowania LAI. Podejście to jest powszechnie określane jako technika inwersji PAR. Rzeczywisty świat jest nieco bardziej złożony, ale jak zobaczysz w sekcji 3, prawo Beera jest podstawą do oszacowania LAI przy użyciu pomiarów padającego i transmitowanego PAR.
Zalety: Technika inwersji PAR jest nieniszcząca, co jest oczywistą, ale główną zaletą, która pozwala na wielokrotne pobieranie próbek z czasoprzestrzeni. Technika inwersji PAR jest również atrakcyjna, ponieważ ma solidne podstawy w teorii transferu promieniowania i biofizyce i ma zastosowanie w wielu różnych typach koron drzew. Z tych powodów technika inwersji PAR jest obecnie standardową i dobrze przyjętą procedurą.
Oprócz przyrządów ręcznych, takich jak ceptometr METER LP-80 ceptometr, standardowe czujniki PAR (tzw. czujniki kwantowe) mogą być również używane do pomiaru transmitowanego promieniowania dla modelu konwersji PAR. Zaletą korzystania z czujników PAR w przeciwieństwie do specjalnie skonstruowanego, ręcznego przyrządu LAI jest to, że czujniki PAR można pozostawić w terenie, aby stale mierzyć zmiany transmitancji PAR. Może to być przydatne podczas badania szybkich zmian LAI w koronach drzew lub gdy nie jest możliwe odwiedzanie terenu wystarczająco często, aby uchwycić czasową zmienność LAI za pomocą ręcznego przyrządu.
Ograniczenia: Technika inwersji PAR ma kilka ograniczeń. Wymaga ona pomiarów zarówno transmitowanego (pod okapem), jak i padającego (nad okapem) PAR w identycznych lub bardzo podobnych warunkach oświetleniowych. Może to stanowić wyzwanie w bardzo wysokich koronach drzew, chociaż pomiary PAR padającego mogą być wykonywane w dużych lukach lub polanach. Ponadto w wyjątkowo gęstych koronach pochłanianie PAR może być prawie całkowite, pozostawiając niewiele światła przechodzącego do pomiaru w dolnej części korony. Utrudnia to rozróżnienie zmian lub różnic w LAI, gdy LAI jest bardzo wysoki. Wreszcie, na szacunki LAI uzyskane z pomiarów transmitowanego PAR mogą mieć wpływ kępy liści. Błędy w szacowaniu LAI związane z kępami mogą być zwykle złagodzone poprzez zbieranie licznych przestrzennie rozmieszczonych próbek transmitowanego PAR.
Współczynnik odbicia promieniowania
Inna metoda szacowania LAI wykorzystuje światło odbite, a nie przechodzące. Promieniowanie odbite od zielonej, zdrowej roślinności ma bardzo wyraźne widmo (rysunek 3). W rzeczywistości niektórzy naukowcy zaproponowali znalezienie potencjalnie nadających się do zamieszkania planet poza naszym Układem Słonecznym, szukając tego unikalnego sygnału widmowego. Typowe widmo odbicia roślinności ma bardzo niski współczynnik odbicia w widzialnej części widma elektromagnetycznego (~400 do 700 nm, co jest również regionem PAR). Jednak w obszarze bliskiej podczerwieni (NIR) (> 700 nm) współczynnik odbicia może wynosić nawet 50%. Dokładna wartość współczynnika odbicia dla każdej długości fali zależy od stężenia różnych pigmentów liściowych, takich jak chlorofil i struktura korony (np. układ i liczba warstw liści).
Zalety: Wczesne próby wykorzystania danych odbicia spektralnego do ilościowego określenia właściwości korony drzew wykazały, że stosunek odbicia czerwonego i NIR może być wykorzystany do oszacowania procentowego pokrycia korony drzew dla danego obszaru. Późniejsze wysiłki doprowadziły do powstania szeregu różnych kombinacji długości fal, które odnoszą się do różnych właściwości koron drzew. Te kombinacje długości fal lub spektralne wskaźniki roślinności są obecnie rutynowo stosowane jako wskaźniki zastępcze dla LAI lub, poprzez modelowanie empiryczne, są wykorzystywane do bezpośredniego szacowania LAI.
Do niedawna jednym z jedynych sposobów gromadzenia danych dotyczących współczynnika odbicia był ręczny spektrometr - drogi, delikatny instrument przeznaczony do laboratorium, a nie do pracy w terenie. Opcje czujników zostały jednak rozszerzone wraz z opracowaniem lekkich radiometrów wielopasmowych, które mierzą określony wskaźnik roślinności. Te małe czujniki są niedrogie i nie wymagają dużej mocy, co czyni je idealnymi do monitorowania w terenie.
To dobra wiadomość dla każdego, kto chce monitorować zmiany LAI w czasie, w tym badaczy zainteresowanych fenologią, wzrostem korony, wykrywaniem stresu i spadku korony lub wykrywaniem chorych roślin.
Wskaźniki roślinności mają jeszcze jedną zaletę: wiele satelitów obserwujących Ziemię, takich jak Quickbird, Landsat i MODIS, mierzy współczynnik odbicia, który można wykorzystać do obliczenia wskaźników roślinności. Ponieważ satelity te obserwują duże obszary, mogą służyć jako sposób skalowania obserwacji dokonanych w skali lokalnej na znacznie szersze obszary. I odwrotnie, pomiary wykonane w skali lokalnej za pomocą radiometru wielopasmowego mogą być użytecznym źródłem danych naziemnych dla satelitarnych wskaźników roślinności.
Radiometry wielopasmowe oferują również opcję top-down dla bardzo krótkich zadaszeń, takich jak krótka trawa preriowa i zarośla. Zastosowanie większości metod szacowania LAI jest trudne, jeśli nie niemożliwe, ponieważ sprzęt jest zbyt duży, aby w pełni zmieścić się pod okapem. Wskaźniki roślinności są mierzone za pomocą czujników, które obserwują koronę z góry na dół, co czyni je doskonałą alternatywą w takich przypadkach.
Rysunek 3. Widma odbicia uzyskane na różnych etapach rozwoju korony. Uwaga: Istnieje wyraźna różnica między współczynnikiem odbicia światła widzialnego i bliskiej podczerwieni (NIR), która rozwija się wraz ze wzrostem LAI.
Ograniczenia: Jednym z największych ograniczeń indeksów roślinności jest to, że są one wartościami bezjednostkowymi i gdy są używane samodzielnie, nie zapewniają bezwzględnej miary leaf area index. Jeśli nie potrzebujesz bezwzględnych wartości LAI, wartość indeksu roślinności może być użyta jako wskaźnik zastępczy dla LAI. Jeśli jednak potrzebujesz bezwzględnych wartości LAI, będziesz musiał użyć innej metody pomiaru LAI w połączeniu z indeksem roślinności, dopóki nie zostanie zebrana wystarczająca ilość danych kolokowanych do stworzenia modelu empirycznego. Metoda ta może być również ograniczona ze względu na lokalizację czujników. Z natury współczynnik odbicia musi być mierzony od szczytu korony rośliny, co może być niewykonalne w przypadku niektórych wysokich korony.
Korzystanie z ceptometru LP-80
Ceptometr METER LP-80 wykorzystuje technikę inwersji PAR do obliczania leaf area index (LAI). Strona LP-80 wykorzystuje zmodyfikowaną wersję modelu transmisji i rozpraszania światła przez korony drzew opracowanego przez Normana i Jarvisa (1975). Poniżej omówiono pięć kluczowych zmiennych wykorzystywanych jako dane wejściowe.
τ (stosunek transmitowanego i padającego PAR): Najbardziej istotnym czynnikiem przy określaniu LAI za pomocą dowolnego modelu inwersji PAR jest stosunek transmitowanego PAR do padającego PAR. Stosunek ten(τ) jest obliczany na podstawie pomiarów transmitowanego PAR w pobliżu powierzchni gruntu i padającego PAR nad okapem.
τ jest stosunkowo intuicyjną zmienną do zrozumienia. Gdy LAI jest niski, większość padającego promieniowania jest transmitowana przez baldachim, a nie pochłaniana lub odbijana, a zatem τ będzie bliskie 1. Wraz ze wzrostem ilości liści w baldachimie następuje proporcjonalny wzrost ilości pochłanianego światła, a malejąca część światła będzie transmitowana do powierzchni ziemi. Strona LP-80 składa się z listwy świetlnej, która ma 80 liniowo rozmieszczonych czujników PAR i zewnętrznej strony PAR sensor. W typowych scenariuszach listwa świetlna służy do pomiaru PAR pod okapem, podczas gdy czujnik zewnętrzny ma na celu ilościowe określenie PAR padającego nad okapem lub na polanie.
θ (kąt zenitalny słońca):θ to kątowa wysokość słońca na niebie w odniesieniu do zenitu lub punktu bezpośrednio nad głową, w danym czasie, dniu i położeniu geograficznym (Rysunek 4). Kąt zenitalny słońca jest używany do opisania długości ścieżki fotonów przez baldachim (np. w zamkniętym baldachimie długość ścieżki wzrasta, gdy słońce zbliża się do horyzontu) oraz do określenia interakcji między promieniowaniem wiązki a orientacją liści (omówioną poniżej).
θ jest automatycznie obliczane przez LP-80 przy użyciu danych wejściowych czasu lokalnego, daty, szerokości i długości geograficznej. Dlatego bardzo ważne jest, aby upewnić się, że są one prawidłowo ustawione w menu konfiguracji LP-80 .
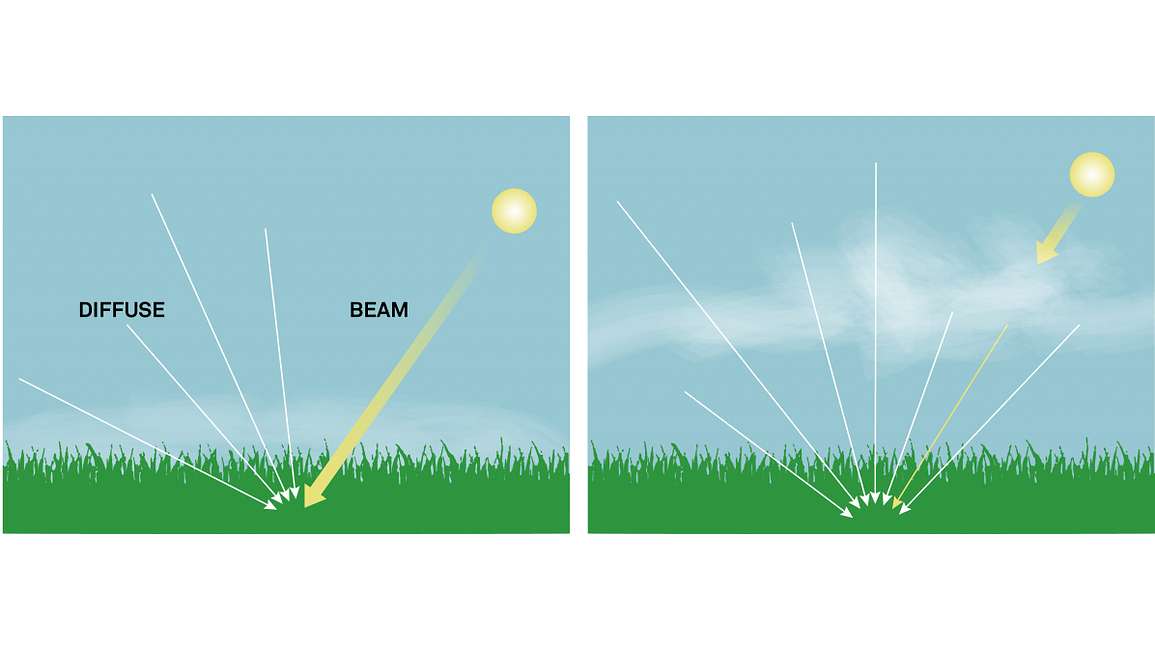
ƒb (frakcja wiązki): W środowisku zewnętrznym ostatecznym źródłem promieniowania krótkofalowego jest słońce. Gdy niebo jest czyste, większość promieniowania pochodzi bezpośrednio od słońca (rysunek 5a). Jednak w obecności chmur lub zamglenia pewna część promieniowania wiązki jest rozpraszana przez parę wodną i aerozole w atmosferze (rysunek 5b). Ten rozproszony składnik jest określany jako promieniowanie rozproszone. ƒb jest obliczane jako stosunek promieniowania rozproszonego do promieniowania wiązki. Strona LP-80 automatycznie oblicza ƒb, porównując zmierzone wartości padającego PAR ze stałą słoneczną, która jest znaną wartością energii świetlnej ze słońca (przy założeniu bezchmurnego nieba) w danym czasie i miejscu na powierzchni ziemi.
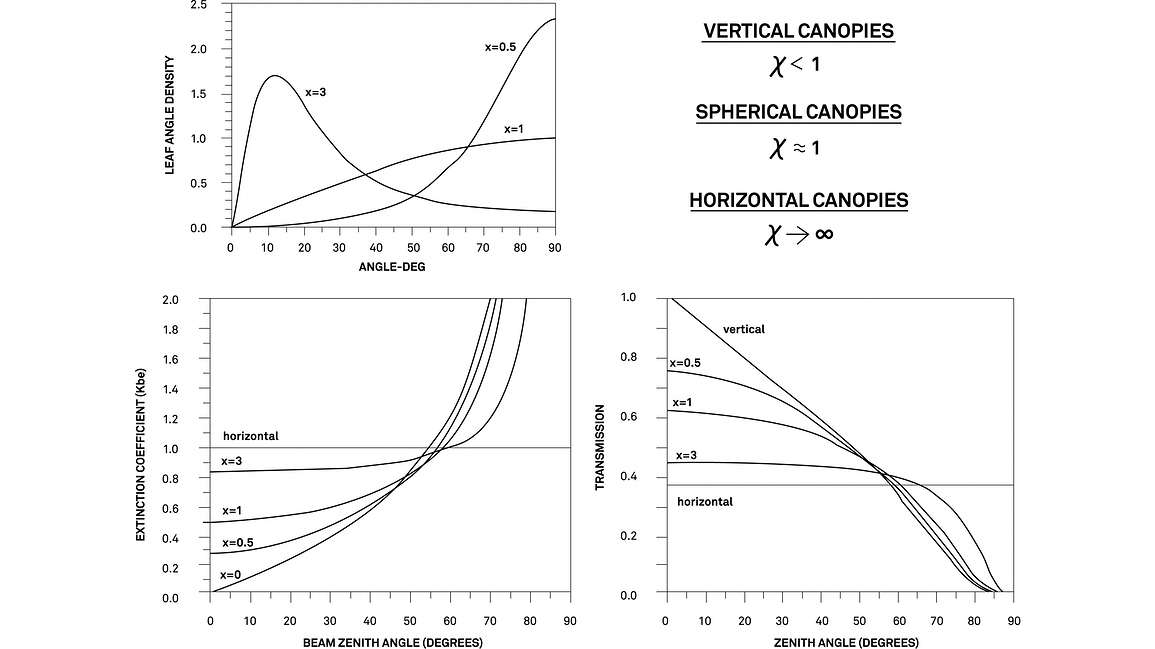
χ (leaf angle distribution): The leaf angle distribution parameter (χ) describes the projection of leaf area onto a surface. Imagine, for example, a light source directly overhead. The shadow cast by a leaf with a vertical orientation would be much smaller than the shadow cast by a leaf with a horizontal orientation. In nature, canopies are typically composed of leaves with a mixture of orientations. This mixture is often best described by what is known as the spherical leaf distribution with a χ value = 1 (the default in the LP-80). Canopies with predominantly horizontal orientations, such as strawberries, have χ values > 1, whereas canopies with predominantly vertical orientations, like some grasses, have χ values < 1.
Ogólnie rzecz biorąc, χ opisuje, ile światła zostanie pochłonięte przez liście w czaszy o różnych porach dnia, gdy słońce przesuwa się po niebie. Oszacowanie leaf area index za pomocą techniki inwersji PAR nie jest zbyt wrażliwe na wartość χ, zwłaszcza podczas pobierania próbek w warunkach równomiernie rozproszonego nieba (Garrigues i in., 2008). Wartość χ jest najważniejsza podczas pracy z zadaszeniami o bardzo pionowej lub poziomej charakterystyce oraz podczas pracy w warunkach czystego nieba, gdzie fb jest mniejsze niż około 0,4. Dodatkowe informacje na temat rozkładu kąta nachylenia liści można znaleźć w Campbell i Norman (1998).
Rysunek 4. Kąty zenitalne słońca zmieniają się w ciągu dnia. Obserwator jest zwrócony w stronę równika.Rysunek 5. Ułamek wiązki w warunkach (a) słonecznego i (b) zachmurzonego nieba
K (współczynnik ekstynkcji): Współczynnik ekstynkcji baldachimu, K, opisuje ilość promieniowania pochłanianego przez baldachim przy danym kącie zenitalnym słońca i rozkładzie kąta nachylenia liści baldachimu. Pojęcie współczynnika ekstynkcji pochodzi z prawa Beera (równanie 1). Szczegółowe wyjaśnienie współczynnika ekstynkcji może szybko stać się skomplikowane. Do oszacowania LAI wystarczy wiedzieć, że kąt penetracji wiązki słonecznej oddziałuje z rozkładem kąta liści, aby określić prawdopodobieństwo, że foton zostanie przechwycony przez liść. Dla celów oszacowania LAI, K jest obliczane jako
Równanie 2
Z tego równania powinno być oczywiste, że dla każdego zadaszenia, K zmienia się tylko wtedy, gdy słońce przesuwa się po niebie. Strona LP-80 automatycznie oblicza K za każdym razem, gdy mierzy LAI. Po obliczeniu K i określeniu wszystkich innych zmiennych, LAI jest obliczany jako
Równanie 3
gdzie L to LAI, a A to chłonność liści. Domyślnie A jest ustawione na 0,9 w LP-80. Chłonność liści jest bardzo spójną właściwością dla większości zdrowych zielonych liści, a wartość 0,9 jest dobrym przybliżeniem dla większości sytuacji. W skrajnych przypadkach (np. bardzo młode liście, liście silnie owłosione lub woskowe, liście starzejące się), A może odbiegać od 0,9, prowadząc do błędów w szacunkach LAI. Jeśli korzystasz z LP-80 w nietypowych warunkach, może być konieczne ręczne połączenie danych wyjściowych z LP-80 ze zmodyfikowaną wartością A w celu obliczenia LAI.
Korzystanie z LP-80 w krótkich zadaszeniach (uprawy zbóż, użytki zielone)
W typowych sytuacjach najlepiej jest trzymać ceptometr LP-80 na stałej wysokości pod okapem, podczas gdy dołączony zewnętrzny PAR sensor jest trzymany nad okapem. Użyj dołączonej poziomicy bąbelkowej, aby upewnić się, że pasek świetlny i zewnętrzny PAR sensor są utrzymywane na poziomie. W przypadku upraw rzędowych lub małych poletek próbnych badacze często montują czujnik zewnętrzny na statywie między rzędami lub nad okapem. Czujnik LP-80 wykonuje jednoczesne pomiary PAR nad i pod okapem za każdym naciśnięciem przycisku, uwzględniając wszelkie zmiany warunków oświetleniowych. Jeśli baldachim jest wystarczająco krótki, jeszcze łatwiejszym podejściem jest użycie ceptometru do uzyskania pomiarów zarówno nad, jak i pod baldachimem. Wystarczy przytrzymać LP-80 nad baldachimem, aby uzyskać pomiar padającego PAR. Pomiar nad okapem należy aktualizować co kilka minut lub w miarę zmiany warunków na niebie (np. z powodu zmiennego zachmurzenia). W obu przypadkach wszystkie inne zmienne są mierzone i obliczane automatycznie, a leaf area index (LAI) jest aktualizowany przy każdym pomiarze pod okapem.
Korzystanie z LP-80 w wysokich zadaszeniach (lasy, obszary nadbrzeżne)
W wysokich zadaszeniach pomiar PAR powyżej i poniżej zadaszenia za pomocą jednego przyrządu jest często niepraktyczny. Podczas korzystania z LP-80 w wysokich zadaszeniach dostępnych jest kilka opcji wykonywania pomiarów PAR powyżej i poniżej zadaszenia.
Jedną z opcji jest zamontowanie kamery PAR sensor nad baldachimem lub na rozległej polanie z niezakłóconym widokiem na niebo. Ta metoda wymaga dodatkowego przetwarzania danych, ale może dać dobre wyniki. Urządzenie PAR sensor musi być podłączone do własnego rejestratora danych, który powinien być skonfigurowany do pozyskiwania pomiarów w regularnych odstępach czasu (np. co 1 do 5 minut), aby uchwycić wszelkie zmiany poziomu oświetlenia otoczenia. Zbierz pomiary pod okapem za pomocą ceptometru, a następnie połącz dane w procesie przetwarzania końcowego, używając znaczników czasu do sparowania każdego pomiaru nad i pod okapem. Oblicz τ dla każdej pary, którą można następnie wykorzystać jako dane wejściowe do równania 3.
Druga opcja jest przydatna, gdy nie jest możliwe umieszczenie urządzenia PAR sensor nad okapem lub gdy urządzenie PAR sensor lub rejestrator danych nie są dostępne. W takim przypadku należy użyć LP-80 do pomiaru padającego PAR w miejscu poza okapem z niezakłóconym widokiem na niebo. W trybie pomiaru należy wybrać pomiar promieniowania padającego lub wysyłanego. Podczas korzystania z samego urządzenia LP-80 do wykonywania odczytów nad i pod okapem należy wziąć pod uwagę zmienność warunków na niebie.
Przy bezchmurnym niebie najłatwiej jest pobierać próbki w środku dnia, ponieważ poziomy światła nie zmieniają się zbytnio w ciągu 20 do 30 minut. Gdy niebo jest jednolicie zachmurzone, warunki PAR mogą utrzymywać się przez dłuższy czas, dając dłuższe okno pomiarowe przed koniecznością ponownego wykonania pomiaru nad okapem.
Jeśli jednak warunki na niebie są bardzo zmienne, nie zalecamy tej metody, chyba że możliwe jest ciągłe aktualizowanie pomiarów PAR. Urządzenie LP-80 automatycznie oblicza LAI przy każdym pomiarze poniżej okapu, korzystając z zapisanego pomiaru PAR. Aby zapobiec błędom w obliczeniach LAI, należy ponownie pobrać pomiar PAR za każdym razem, gdy zmienią się warunki oświetleniowe (np. gdy cloud zasłania tarczę słoneczną lub po upływie około 20-30 minut).
Grupowanie i próbkowanie przestrzenne
W większości zadaszeń, leaf area index jest zmienny w przestrzeni. Na przykład w uprawach rzędowych LAI może wahać się od 0 do 2-3 w odległości 1 metra. Nawet w lasach i innych naturalnych zadaszeniach, zmienne odstępy między drzewami, charakterystyka rozgałęzień i układ liści na łodygach powodują tworzenie się kęp. Oznacza to, że punktowe pomiary LAI mogą być wysoce stronnicze. Lang i Yueqin (1986) odkryli, że uśrednienie kilku pomiarów wzdłuż poziomego transektu pomogło złagodzić odchylenia związane z grupowaniem się w drobnych skalach przestrzennych.
System LP-80 wykorzystuje podobne podejście, uśredniając pomiary światła w ośmiu grupach po dziesięć czujników rozmieszczonych wzdłuż sondy o długości 80 cm. Chociaż podejście to zmniejsza błędy w skali lokalnej, może nie uwzględniać zmienności leaf area index w skali korony. Badacze muszą wziąć pod uwagę przestrzenną zmienność LAI w koronach drzew podczas opracowywania schematu pobierania próbek. Ogólnie rzecz biorąc, bardziej niejednorodne korony drzew będą wymagały większej liczby pomiarów LAI w przestrzeni w celu uzyskania wartości LAI reprezentatywnej dla całej korony.
Warunki atmosferyczne
Urządzenie LP-80 jest w stanie dokładnie mierzyć leaf area index zarówno przy czystym niebie, jak i przy zachmurzeniu. Wynika to z faktu, że model LAI używany przez LP-80 uwzględnia zmiany w promieniowaniu rozproszonym i wiązce(ƒb), kącie zenitalnym słońca(θ), a także dlatego, że promieniowanie padające i przepuszczane są mierzone jednocześnie podczas korzystania z PAR sensor. Błędy związane z nieprawidłowym określeniem rozkładu kąta nachylenia liści(χ) są najbardziej widoczne podczas pobierania próbek w warunkach bezchmurnego nieba (Garrigues i in., 2008). Wynika to z faktu, że większa część promieniowania pochodzi z jednego kąta (promieniowanie wiązki bezpośrednio od słońca). W takich warunkach ważne jest prawidłowe modelowanie interakcji kąta nachylenia liści i kąta penetracji wiązki. Tak więc, podczas pobierania próbek w warunkach bezchmurnego nieba, należy upewnić się, że używana jest odpowiednia wartość χ.
Wpływ elementów niefotosyntetycznych
W lasach, krzewach i innych obszarach, w których występują gatunki drzewiaste, na pomiary LP-80 będą miały wpływ elementy inne niż liście. Na przykład pnie drzew, gałęzie i pnie będą przechwytywać część promieniowania, a tym samym wpływać na szacunki LAI uzyskane za pomocą techniki inwersji PAR. W rzeczywistości niektórzy badacze odnoszą się do pomiaru uzyskanego z LP-80 i podobnych instrumentów jako wskaźnika powierzchni roślin (PAI), a nie LAI, aby uznać wkład materiału niebędącego liśćmi w pomiar. Nie powinno dziwić, że PAI będzie wyższy niż LAI w danym ekosystemie. Jednak wartości PAI i LAI często nie różnią się zbytnio, ponieważ powierzchnia liści jest na ogół znacznie większa niż powierzchnia gałęzi, a większość gałęzi jest zacieniona przez liście (Kucharik i in., 1998). W ekosystemach liściastych udział materiału drzewnego można uwzględnić, wykonując pomiary w fazie opadania liści.
Korzystanie z czujnika SRS-NDVI
Czujnik SRS-NDVI mierzy współczynnik odbicia korony w czerwonych i podczerwonych długościach fal, co pozwala na obliczenie znormalizowanego wskaźnika różnicowego wegetacji (NDVI). Z kolei NDVI można wykorzystać do oszacowania LAI. Poniżej przedstawiamy krótki przegląd teorii działania SRS-NDVI . SRS-NDVI mierzy współczynnik odbicia światła w koronie w zakresie długości fal czerwonych i NIR, a jego pomiary mogą być wykorzystane do obliczenia lub przybliżenia LAI. Współczynniki odbicia fal czerwonych i NIR są wykorzystywane w następującym równaniu do obliczenia NDVI
Równanie 4
gdzie ρ oznacza procentowy współczynnik odbicia w zakresie długości fal NIR i czerwonych. Matematycznie, NDVI może wahać się od -1 do 1. Wraz ze wzrostem LAI, współczynnik odbicia promieniowania czerwonego zazwyczaj spada ze względu na rosnącą zawartość chlorofilu w koronie, podczas gdy współczynnik odbicia promieniowania NIR wzrasta ze względu na powiększanie się komórek mezofilu i rosnącą złożoność strukturalną korony. Tak więc, w typowych warunkach terenowych, wartości NDVI wahają się od około 0 do 1, reprezentując odpowiednio niski i wysoki LAI.
Rysunek 6. NDVI ściśle śledzi sezonową dynamikę LAI z roku na rok w mieszanym lesie liściastym.
W przypadkach takich jak fenologia i fenotypowanie stay-green, gdzie bezwzględne wartości LAI nie są wymagane, wartości NDVI mogą być używane bezpośrednio jako zamienniki LAI. Na przykład, jeśli celem badania jest śledzenie czasowych wzorców wzrostu i starzenia się korony (Rysunek 6), wówczas odpowiednie może być po prostu użycie NDVI jako metryki. Jeśli cele badawcze wymagają oszacowania rzeczywistego LAI, możliwe jest ustanowienie modelu specyficznego dla okapu, który pozwoli na konwersję NDVI na LAI. Metoda ta została opisana w następnej sekcji.
Opracowanie terenowych modeli regresji NDVI-LAI
Aby bezpośrednio oszacować leaf area index przy użyciu wartości NDVI , należy opracować zależność korelacyjną specyficzną dla danego miejsca lub uprawy. Najlepszym sposobem jest wykonanie kolokowanych pomiarów NDVI i LAI (np. za pomocą ceptometru LP-80 ). Na przykład, kolokowane pomiary LAI i NDVI zostały uzyskane w okresie szybkiego wzrostu korony. Regresja najmniejszych kwadratów została wykorzystana do dopasowania modelu liniowego do danych (Rysunek 7). Dzięki temu modelowi możliwe jest wykorzystanie NDVI do przewidywania LAI bez wykonywania niezależnych pomiarów.
Opracowanie solidnego modelu empirycznego wymaga pewnego wysiłku, ale gdy model jest już gotowy, można stale monitorować zmiany LAI za pomocą czujnika SRS-NDVI rozmieszczonego na działce lub koronie długoterminowo. Metoda ta pozwala zaoszczędzić znaczny wysiłek i czas w dłuższej perspektywie.
Rysunek 7. Związek między NDVI a LAI. Uwaga: dopasowany model regresji liniowej (linia ciągła) można wykorzystać do przewidywania LAI na podstawie pomiarów NDVI .
SRS-NDVI rozważania dotyczące pobierania próbek
Czujnik SRS-NDVI jest przeznaczony do użytku jako czujnik z podwójnym polem widzenia. Oznacza to, że jeden czujnik, posiadający półkuliste pole widzenia, powinien być zamontowany w kierunku nieba. Drugi czujnik, o polu widzenia 36° (kąt połówkowy 18°), powinien być zamontowany w kierunku czaszy. Pomiary w dół i w górę zebrane z każdego czujnika są wykorzystywane do obliczania procentowego współczynnika odbicia w pasmach czerwonym i NIR. Procentowe współczynniki odbicia są używane jako dane wejściowe do równania NDVI (równanie 4).
Czujnik patrzący w górę musi być umieszczony nad wszelkimi przeszkodami, które mogą blokować widok nieba przez czujnik. Czujnik skierowany w dół powinien być skierowany na obszar okapu, który ma być mierzony. Rozmiar obszaru mierzonego przez czujnik skierowany w dół zależy od wysokości czujnika nad okapem. Średnica plamki czujnika skierowanego w dół jest obliczana jako
Równanie 5
gdzie γ to kąt połowy pola widzenia (18° dla SRS-NDVI), a h to wysokość czujnika nad okapem. Dotyczy to pomiaru średnicy plamki, gdy czujnik skierowany w dół jest skierowany prosto w dół (tj. kąt widzenia nadir). W przypadkach, gdy czujnik skierowany w dół jest skierowany poza nadir, plamka będzie ukośna i będzie większa niż obliczona za pomocą równania 5.
Aby określić zmienność przestrzenną LAI, można skonfigurować kilka czujników skierowanych w dół, aby monitorować różne części okapu. Na przykład, kilka czujników zostało zamontowanych nad koronami drzew w lesie liściastym w celu monitorowania różnic w fenologii wiosennej kilku drzew. Pomiary NDVI ujawniły różnice w czasie i wielkości wzrostu liści między drzewami, które zostały zmierzone (Rysunek 8). Podobne podejście można zastosować do monitorowania reakcji roślin na poszczególnych poletkach poddanych eksperymentalnej manipulacji lub do monitorowania wzorców wzrostu w różnych jednostkach rolniczych.
Rysunek 8. Przestrzenna zmienność NDVI podczas wiosennego zazieleniania. Uwaga: Zmienność wynika z różnic w czasie rozwoju liści między drzewami i gatunkami drzew.
Wpływ tła gleby na pomiary NDVI
Znaczny błąd w pomiarach NDVI może wystąpić, gdy gleba znajduje się w polu widzenia czujnika SRS-NDVI lub w sytuacjach, gdy ilość gleby w polu widzenia zmienia się z powodu wzrostu korony (np. od wczesnego do późnego sezonu wegetacyjnego). Qi et al. (1994) wykazali, że NDVI jest wrażliwy zarówno na teksturę gleby, jak i jej wilgotność. Ta wrażliwość gleby może utrudnić porównanie wartości NDVI zebranych w różnych lokalizacjach lub w różnych porach roku. Może to również utrudnić stworzenie wiarygodnego modelu regresji NDVI-LAI. Zmodyfikowany wskaźnik roślinności dostosowany do gleby (MSAVI) został opracowany przez Qi i in. (1994) jako wskaźnik roślinności, który ma niewielką lub żadną wrażliwość na glebę. MSAVI jest obliczany jako
Zalety MSAVI obejmują: (1) nie wymaga regulacji parametrów gleby oraz (2) wykorzystuje dokładnie te same dane wejściowe, co NDVI (współczynniki odbicia światła czerwonego i NIR), co oznacza, że można je obliczyć na podstawie danych wyjściowych dowolnego czujnika NDVI .
Rysunek 9. NDVI ma ograniczony sens dla wartości leaf area index (LAI) większych niż 3 do 4.
Radzenie sobie z nasyceniem NDVI w czaszach o wysokim LAI
Oprócz wrażliwości na glebę, NDVI cierpi również na brak wrażliwości na zmiany LAI, gdy LAI jest większy niż około 3 do 4, w zależności od korony (Rysunek 9). Zmniejszona wrażliwość NDVI przy wysokim LAI wynika z faktu, że chlorofil jest wysoce wydajnym pochłaniaczem promieniowania czerwonego. Dlatego w pewnym momencie dodanie większej ilości chlorofilu do łanu (np. poprzez dodanie materiału liściowego) nie zmieni znacząco współczynnika odbicia promieniowania czerwonego (patrz rysunek 3).
Opracowano kilka rozwiązań dla NDVI nasycenia. Jedno z najprostszych rozwiązań wykorzystuje współczynnik wagowy, który jest stosowany do współczynnika odbicia w bliskiej podczerwieni zarówno w liczniku, jak i mianowniku równania 4. Wynikowy wskaźnik nazywany jest wskaźnikiem roślinności o szerokim zakresie dynamiki (WDRVI; Gitelson, 2004). Współczynnik ważenia może być dowolną liczbą od 0 do 1. Gdy współczynnik ważenia zbliża się do 0, liniowość korelacji WDRVI-LAI ma tendencję do zwiększania się kosztem zmniejszenia wrażliwości na zmiany LAI w rzadkich koronach drzew.
Enhanced Vegetation Index (EVI) to kolejny wskaźnik roślinności, który ma wyższą czułość na wysoki LAI w porównaniu do NDVI. EVI został pierwotnie zaprojektowany do pomiaru z satelitów i zawierał niebieskie pasmo jako dane wejściowe, aby złagodzić problemy związane z patrzeniem przez atmosferę na powierzchnię ziemi z orbity. Niedawno opracowano nową formułę EVI, która nie wymaga niebieskiego pasma. Ta zmodyfikowana wersja EVI jest określana jako EVI2 (Jiang et al., 2008). Podobnie jak indeks MSAVI, EVI2 wykorzystuje dokładnie te same dane wejściowe co NDVI (odbicia w paśmie czerwonym i NIR) i jest obliczany jako
Równanie 7
Kolejną zaletą EVI2 jest mniejsza wrażliwość na glebę w porównaniu do NDVI. EVI2 jest zatem dobrym wszechstronnym wskaźnikiem roślinności do szacowania LAI, ponieważ ma niską wrażliwość na glebę i ma liniową zależność z LAI.
Dowiedz się więcej o NDVI
W poniższym webinarium dr Steve Garrity omawia teorię NDVI i PRI, metody, ograniczenia, zastosowania i nie tylko. Wyjaśnia również czujniki odbicia spektralnego i ich rozważania pomiarowe.
Szybki wykres porównawczy metod LAI
*Labor intensive
**Single with LP-80Continuous with subcanopy PAR sensors
*Requires access to top of canopy or large open area**Requires access to top of canopy
Metoda
Koszt względny
Próbkowanie czasowe
Przydatność dla wysokich zadaszeń
Przydatność dla krótkich zadaszeń
Skalowanie przestrzenne
Łatwość pobierania próbek
Próbki profilowania pionowego
Niszczycielskie zbiory
H*
Pojedynczy
L
H
L
VL
Tak
Pułapki na śmieci
M*
Pojedynczy
H
L
L - M
M
Nie
Fotografia półkulista
M
Pojedynczy
H
L
M
M
Nie
Inwersja PAR (LP-80)
M
Oba*
H*
H
M
H
Tak
Wskaźnik roślinności
L - VH
Ciągłe
M**
VH
M -H
VH
Nie
Tabela 1.KLUCZ:VL = bardzo niski, L = niski, M = umiarkowany, H = wysoki, VH = bardzo wysoki
Środowisko pracy: 0 do 50°C, 0 do 100% wilgotności względnej
Długość sondy: 86,5 cm
Liczba czujników: 80
Długość całkowita: 102 cm (40,25 cala)
Wymiary mikrokontrolera: 15,8 x 9,5 x 3,3 cm (6,2 x 3,75 x 1,3 in)
Zakres PAR: 0 do >2 500 µmol m-2 s-1
Rozdzielczość: 1 µmol m-2 s-1
Minimalna rozdzielczość przestrzenna: 1 cm
Pojemność pamięci danych: 1 MB RAM, 9000 odczytów
Interwał rejestrowania nienadzorowanego: Wybierany przez użytkownika, od 1 do 60 minut
Waga urządzenia: 1,22 kg (2,7 funta)
Pobieranie danych: Bezpośrednio przez kabel RS-232
Zasilanie: 4 ogniwa alkaliczne AA
Zewnętrzne złącze PAR sensor : 3-stykowe okrągłe złącze z blokadą (kabel 2 m)
Opcjonalny kabel przedłużający: 7,6 m (25 stóp)
Referencje
Więcej zasobów odpowiadających na pytania: Czym jest leaf area index i jak mierzyć leaf area index.
Campbell, Gaylon S. i John M. Norman. "Środowisko świetlne baldachimów roślinnych". W An Introduction to Environmental Biophysics, s. 247-278. Springer New York, 1998.
Garrigues, Sébastien, N. V. Shabanov, K. Swanson, J. T. Morisette, F. Baret i R. B. Myneni. "Intercomparison and sensitivity analysis of Leaf Area Index retrievals from LAI-2000, AccuPAR, and digital hemispherical photography over croplands." Agricultural and Forest Meteorology 148, nr 8 (2008): 1193-1209.
Gitelson, Anatoly A. "Wide dynamic range vegetation index for remote quantification of biophysical characteristics of vegetation." Journal of Plant Physiology 161, nr 2 (2004): 165-173.
Hyer, Edward J., i Scott J. Goetz. "Porównanie i analiza wrażliwości instrumentów i metod radiometrycznych do szacowania LAI: oceny z obszaru lasu borealnego". Agricultural and Forest Meteorology 122, no. 3 (2004): 157-174.
Jiang, Zhangyan, Alfredo R. Huete, Kamel Didan i Tomoaki Miura. "Development of a two-band enhanced vegetation index without a blue band." Remote Sensing of Environment 112, nr 10 (2008): 3833-3845.
Kucharik, Christopher J., John M. Norman i Stith T. Gower. "Pomiary powierzchni gałęzi i dostosowanie pomiarów pośrednich leaf area index ". Agricultural and Forest Meteorology 91, nr 1 (1998): 69-88.
Lang, A. R. G. i Xiang Yueqin. "Szacowanie leaf area index na podstawie transmisji bezpośredniego światła słonecznego w nieciągłych zadaszeniach". Agricultural and Forest Meteorology 37, no. 3 (1986): 229-243.
Norman, J. M. i P. G. Jarvis. "Photosynthesis in Sitka spruce (Picea sitchensis (Bong.) Carr.). III. Pomiary struktury korony i przechwytywania promieniowania". Journal of Applied Ecology (1974): 375-398.
Rouse Jr, J_W, R. H. Haas, J. A. Schell i D. W. Deering. "Monitorowanie systemów wegetacyjnych na Wielkich Równinach za pomocą ERTS". (1974).
Qi, Jiaguo, Abdelghani Chehbouni, A. R. Huete, Y. H. Kerr i Soroosh Sorooshian. "Zmodyfikowany indeks roślinności dostosowany do gleby". Remote Sensing of Environment 48, nr 2 (1994): 119-126.
The LP-80: bez bólu leaf area index
DR. GAYLON S. CAMPBELL
Leaf area index (LAI) to tylko pojedyncza liczba - statystyczna migawka korony drzewa wykonana w określonym czasie. Ale ta jedna liczba może prowadzić do znaczącego wglądu, ponieważ może być wykorzystana do modelowania i zrozumienia kluczowych procesów korony, w tym przechwytywania promieniowania, konwersji energii, pędu, wymiany gazowej, przechwytywania opadów i ewapotranspiracji.
Starsze metody LAI są żmudne
Leaf area index jest definiowana jako jednostronna powierzchnia zielonych liści w łanie lub zbiorowisku roślin na jednostkę powierzchni gruntu. Można ją określić poprzez zebranie i zmierzenie powierzchni każdego liścia w łanie pokrywającym jednostkę powierzchni gruntu. W 1981 roku Anderson opracował mniej destrukcyjną metodę określania LAI. Korzystając z półkulistych zdjęć patrzących w górę, oszacowała ułamek światła, który przeniknął przez koronę i zastosowała predykcyjny model matematyczny do przybliżenia leaf area index.
Ocenianie zdjęć czasz typu "rybie oko" było żmudną pracą. Asystent zwykle kładł siatkę na każdym zdjęciu i liczył, jaka część kwadratów była jasna. Jeden z techników laboratoryjnych wspomina: "Po zbyt wielu godzinach oglądania tych zdjęć marzyłem o grze w warcaby". Ocena "warcabów" pozwoliła badaczom znaleźć prawdopodobieństwo, że losowa wiązka światła przeniknie przez daną sekcję czaszy.
Konwersja LAI
Uzyskanie wartości dla leaf area index jest często tylko punktem na drodze. Jeśli planujesz wykorzystać LAI do modelowania interakcji środowiskowych czaszy, pomiar fotosyntetycznie aktywnego promieniowania (PAR) może być bardziej bezpośrednią drogą. Dzieje się tak dlatego, że wiele z tych modeli wykorzystuje LAI do przewidywania PAR. Możliwe jest cofnięcie się w drugą stronę - wykorzystanie PAR do oszacowania LAI. Ale po co to robić, jeśli PAR jest liczbą, której naprawdę potrzebujesz? Warto ocenić, czy LAI jest najbardziej użytecznym parametrem dla konkretnego zastosowania. Czasami prostsze i zwykle dokładniejsze jest po prostu zmierzenie przechwyconego PAR i wykorzystanie tych danych bezpośrednio w odpowiednim modelu.
Model matematyczny, który przekształca ten ułamek światła w szacunkową wartość leaf area index jest stosunkowo prosty. Aby zrozumieć, jak to działa, wyobraź sobie, że trzymasz liść o powierzchni dziesięciu centymetrów kwadratowych poziomo nad dużym białym kwadratem. Rzucałby on cień o powierzchni dziesięciu centymetrów kwadratowych. Następnie losowo umieść liść o identycznym rozmiarze nad kwadratem. Z dużym prawdopodobieństwem rzucany cień miałby teraz dwadzieścia centymetrów kwadratowych, choć istnieje niewielka szansa, że liście nałożyłyby się na siebie. Po dodaniu trzeciego liścia prawdopodobieństwo nakładania się wzrasta. W miarę losowego umieszczania coraz większej liczby liści, biały kwadrat zostanie całkowicie zacieniony. I choć powierzchnia liści będzie się zwiększać w miarę ich dodawania, zacieniony obszar pozostanie stały, ponieważ całe światło zostanie przechwycone.
Strona LP-80 rozwiązuje równanie za użytkownika
Równanie opisujące to zjawisko (patrz Rozwiązywanie równania poniżej w celu jego matematycznego wyprowadzenia) to
Równanie 1
τ to prawdopodobieństwo, że promień przeniknie przez baldachim, L to leaf area index baldachimu, a K to współczynnik ekstynkcji baldachimu. Jeśli zmierzysz fotosyntetycznie aktywne promieniowanie zarówno powyżej, jak i poniżej czaszy w jasny słoneczny dzień, stosunek tych dwóch (PAR poniżej do PAR powyżej) jest w przybliżeniu równy τ. Jeśli znasz K, możesz znaleźć leaf area index (L), odwracając równanie:
Równanie 2
Równanie LP-80 zasadniczo rozwiązuje to równanie, aby znaleźć leaf area index. Istnieje jednak kilka czynników komplikujących. Podczas konstruowania modelu założyliśmy, że liście w naszym sztucznym baldachimie są poziome i czarne, a całe promieniowanie pochodzi bezpośrednio od słońca. W rzeczywistości kąt padania promieni słonecznych zmienia się w ciągu dnia, a prawdziwe zadaszenia mają dość złożoną architekturę. Ponadto część promieniowania jest rozpraszana zarówno przez liście w baldachimie, jak i przez niebo. Pełny model obliczania leaf area index na podstawie pomiaru promieniowania fotosyntetycznie czynnego obejmuje poprawki dla wszystkich tych czynników.
Równanie 3
To równanie, które jest faktycznie używane przez LP-80, dostosowuje się do ilości światła pochłoniętego (a nie rozproszonego) przez liście w terminie A oraz do frakcji światła, które wchodzi do czaszy jako wiązka (w przeciwieństwie do rozproszonego światła z nieba lub chmur) w terminie fb. K, współczynnik ekstynkcji czaszy, zawiera zmienne dla kąta zenitalnego słońca i rozmieszczenia liści. Jeśli określisz swoją lokalizację i ustawisz wewnętrzny zegar na czas lokalny, LP-80 obliczy kąt zenitalny słońca w czasie każdego pomiaru. Zakłada się, że rozkład kąta liści jest sferyczny, chyba że użytkownik wskaże inaczej.
Rozwiązanie równania
Jeśli podzielimy baldachim losowo rozmieszczonych poziomych czarnych liści na tak wiele warstw, że każda warstwa zawiera nieskończenie mały ułamek powierzchni liści(dL), zmiana promieniowania od góry do dołu tej warstwy wynosi
Równanie 4
Innymi słowy, zmiana średniej ilości światła słonecznego przechodzącego przez tę część czaszy(dSb) jest równa ujemnej (ponieważ ilość światła zmniejsza się wraz ze wzrostem powierzchni liści) średniej ilości mocy promieniowania na jednostkę powierzchni(Sb) pomnożonej przez zmianę leaf area index (dL). Jest to zmienne rozdzielne równanie różniczkowe. Dzieląc obie strony przez Sb i całkując od szczytu baldachimu w dół, otrzymujemy
Równanie 5
Wykonanie całkowania daje
Równanie 6
Biorąc wykładnik z obu stron otrzymujemy
Równanie 7
Sbo to promieniowanie na poziomej powierzchni nad baldachimem; τ to prawdopodobieństwo, że promień przeniknie przez baldachim, które jest takie samo jak stosunek promieniowania wiązki na dole baldachimu do promieniowania wiązki na górze (ponieważ zakładamy brak rozpraszania promieniowania w baldachimie). W przypadku baldachimów z liśćmi innymi niż poziome, wynik jest taki sam, z wyjątkiem tego, że L jest zastępowane przez KL, gdzie K jest współczynnikiem ekstynkcji baldachimu.
Odniesienie
Anderson, Margaret C. "Geometria rozmieszczenia liści w niektórych lasach południowo-wschodniej Australii". Agricultural Meteorology 25 (1981): 195-206. Link do artykułu.
The LP-80: Jak dokładny jest ten system?
Urządzenie LP-80 umożliwia szybkie, bezpośrednie pomiary promieniowania fotosyntetycznie czynnego(PAR) w zadaszeniach. Zapewnia natychmiastowe pomiary PAR po włączeniu, a także zapewnia pomiar Leaf Area Index-LAI. Ale skąd pochodzi ten pomiar LAI i jak dokładny jest?
Daszki są zmienne. Strona LP-80 uwzględnia to za pomocą χ.
Leaf area index to jednostronna, zielona powierzchnia liści łanu lub zbiorowiska roślin na jednostkę powierzchni gruntu. Aby bezpośrednio zmierzyć LAI, należałoby zmierzyć powierzchnię każdego liścia w łanie powyżej jednostki powierzchni gruntu. Ponieważ metoda ta jest zarówno destrukcyjna, jak i niezwykle czasochłonna, jest rzadko stosowana. Wszystkie inne pomiary wskaźnika powierzchni liści, od zdjęć półkulistych po czujniki optyczne, próbują przybliżyć tę wartość. Strona LP-80 określa LAI poprzez pomiar fotosyntetycznie aktywnego promieniowania i konwersję wartości PAR na leaf area index. LP-80 wykorzystuje kilka zmiennych do obliczenia leaf area index. Jedna z tych zmiennych, χ, opisuje orientację liści w łanie.
Co to jest χ?
χ to "parametr rozkładu kąta nachylenia czaszy". Opisuje on architekturę czaszy - w jaki sposób jej liście są zorientowane w przestrzeni. Mówi się, że liście rozmieszczone losowo w przestrzeni mają rozkład sferyczny, co oznacza, że gdyby każdy liść w czaszy został ostrożnie przesunięty bez zmiany jego orientacji, liście mogłyby zostać wykorzystane do pokrycia powierzchni kuli. Baldachim ze sferycznie rozmieszczonymi liśćmi ma wartość χ równą 1.
Many canopy architectures tend to be more horizontal (χ > 1) or vertical (χ < 1). Some canopy types have published χ values (see the LP-80 manual for a short list). But because this value can vary from species to species, it’s important to be able to approximate the value.
LAI czy PAR: kto pierwszy?
Uzyskanie wartości dla Leaf Area Index jest często tylko punktem na drodze. Jeśli planujesz wykorzystać LAI do modelowania interakcji środowiskowych czaszy, pomiar fotosyntetycznie aktywnego promieniowania (PAR) może być bardziej bezpośrednią drogą. Dzieje się tak, ponieważ wiele modeli matematycznych wykorzystuje LAI do przewidywania PAR w swoich wewnętrznych równaniach. Czasami badacze używają PAR do przewidywania LAI, a następnie nieświadomie umieszczają liczbę LAI w modelu, który działa w drugą stronę. Ważne jest, aby ocenić, czy LAI jest najbardziej użytecznym parametrem w konkretnym zastosowaniu. Czasami prostsze i zazwyczaj dokładniejsze jest po prostu zmierzenie przechwyconego PAR i wykorzystanie tych danych bezpośrednio w odpowiednim modelu.
Dobra wartość χ poprawia dokładność aproksymacji LAI
Kuszące jest poszukiwanie dokładnej liczby dla χ, z dokładnością do co najmniej kilku miejsc po przecinku. Jednak ze względu na niewiarygodną zmienność czasz, tego rodzaju dokładność jest niemożliwa do osiągnięcia. Leaf area index liczby, choć cenne, zawsze są tylko przybliżeniami. Dobra wartość χ poprawia dokładność przybliżenia leaf area index (LAI). Ale nawet przy mniej dokładnej wartości χ, przybliżenia leaf area index będą prawdopodobnie dość dokładne w zależności od innych warunków (patrz rysunek 1).
Aby przybliżyć wartość χ dla okapu, znajdź reprezentatywną kępę okapu o równej głębokości i szerokości. Następnie określ frakcję szczeliny pionowej (τ0) - procent światła do cienia, który widzisz pionowo przez kępę - oraz frakcję szczeliny poziomej (τ90) - procent światła, który widzisz poziomo przez kępę. Na przykład w baldachimie z idealnie pionowymi liśćmi można zobaczyć około 10% światła do 90% cienia w poziomie - (τ90) = 0,1 i 100% światła w pionie - (τ0) = 1. χ można znaleźć z następującego prostego równania
Równanie 1
Używając tego równania, χ = 0 dla idealnie pionowego baldachimu. Jeśli liście byłyby rozmieszczone sferycznie, a około 10% światła byłoby widoczne zarówno w pionie, jak i w poziomie, (τ90) = (τ0) = 0,1. Następnie użyj tego równania, χ = 1. (Jest to, nawiasem mówiąc, domyślne ustawienie χ na stronie LP-80).
Minimalizacja niepewności ostatecznej wartości LAI
Ze względów praktycznych oszacowanie ilości światła widocznego przez "reprezentatywną kępę" okapu może być trudne. Łatwiejsze może okazać się wykonanie tła i wykorzystanie go do analizy okapu (my użyliśmy kwadratu o wymiarach 1 metr na 1 metr wykonanego z kolorowej tablicy plakatowej). Kępa powinna zawierać wszystkie typowe elementy łanu. Jeśli na przykład badasz uprawy rzędowe, kępa powinna rozciągać się od środka jednego rzędu do środka następnego, aby uwzględnić charakterystyczną lukę w łanie, która występuje między rzędami. Wyobraź sobie rozcięcie kępy na sześcian. Aby oszacować τ, użyj tła, aby utworzyć tylną stronę sześcianu i ustaw się z przodu, aby oszacować procent światła przepuszczanego poziomo przez tę sześcienną część baldachimu. Aby oszacować τ0, użyj tła, aby utworzyć górną lub dolną część sześcianu i ustaw się na przeciwległym końcu, aby oszacować procent światła przepuszczanego w pionie. Następnie znajdź χ z równania 1 (pokazanego powyżej).
Sprawdź zasadność swoich szacunków, pamiętając, że wartości χ dla bardziej poziomych okapów są większe niż jeden, podczas gdy wartości χ dla bardziej pionowych okapów są mniejsze niż jeden. Wartość χ dla okapu można określić, wybierając opcję "Set χ" w menu Setup (Konfiguracja) aplikacji LP-80. Korzystając z tej metody, powinieneś być w stanie oszacować wartość χ, która zminimalizuje niepewność ostatecznej wartości leaf area index .
Rysunek 1. Procentowy błąd w obliczeniach L na stronie LP-80 , jeśli LP-80 jest ustawiony na ?=1, a rzeczywisty parametr rozkładu okapu ma wartość pokazaną na rysunku.
Ten rysunek przedstawia procentowy błąd w obliczeniach L na stronie LP-80 , jeśli LP-80 jest ustawiony na ? = 1, a rzeczywisty parametr rozkładu okapu ma wartość pokazaną na rysunku. Zakłada się pełne nasłonecznienie (fb= 0,8). Należy pamiętać, że błąd zależy od kąta zenitalnego słońca. Większość pomiarów będzie wykonywana przy kątach zenitalnych większych niż 30 stopni, więc błąd w pełnym słońcu, bez informacji o parametrze rozkładu okapu, wynosi w najgorszym przypadku 20%. Błąd ten zmniejsza się wraz ze spadkiem wartości fb i wynosi zero, gdy fb wynosi zero. Jeśli parametr rozkładu koron drzew można oszacować z dokładnością 10% lub lepszą, błąd LAI wyniesie 5% lub więcej nawet przy zerowym kącie zenitalnym. W związku z tym niepewność parametru dystrybucji prawdopodobnie nie przyczyni się znacząco do niepewności LAI.
DODATEK
Uproszczone modele asymilacji węgla przez rośliny
Dr Gaylon S. Campbell
Szczegółowe procesy fotosyntezy są skomplikowane i trudne do modelowania. W wielu przypadkach możliwe jest jednak uproszczenie modelu poprzez skupienie się na jednym lub kilku ograniczeniach asymilacji.
Uproszczona asymilacja węgla: światło i woda
Mówiąc najprościej, asymilacja węgla obejmuje chemiczną przemianę dwutlenku węgla i wody w węglowodany i tlen w liściach roślin. Proces ten wymaga energii, która jest dostarczana przez światło, zwykle pochodzące ze słońca.CO2 pochodzi z atmosfery i musi dyfundować do komórek mezofilu liścia, aby ulec wiązaniu. Ponieważ wnętrze liścia jest znacznie bardziej wilgotne niż atmosfera, woda dyfunduje na zewnątrz, gdyCO2 dyfunduje do środka. Ilość wody zużywanej w rzeczywistym procesie fotosyntezy jest niewielka, ale woda tracona w związku z pobieraniemCO2 jest znaczna.
Ograniczone przez światło, ograniczone przez wodę: dwa różne podejścia
Opierając się na tym prostym opisie, moglibyśmy postulować sytuacje, w których światło byłoby czynnikiem ograniczającym asymilację, a inne, w których czynnikiem ograniczającym byłaby woda. Nasze modele mogą wyglądać następująco: asymilacja jest proporcjonalna do zdolności rośliny do wychwytywania światła lub asymilacja jest proporcjonalna do zdolności rośliny do wychwytywania wody. Oba podejścia mogą być przydatne w modelowaniu produkcji biomasy.
Model oparty na świetle
Model oparty na świetle w postaci równania to
Równanie 1
gdzie A jest asymilacją netto suchej masy, S jest całkowitym promieniowaniem padającym otrzymanym w czasie wzrostu uprawy, f jest średnią frakcją promieniowania przechwyconego przez uprawę, a e jest wydajnością konwersji. Jeśli A i S są wyrażone w mol m-2s-1, to e jest bezwymiarową wydajnością konwersji. W sytuacjach ograniczających dostęp światła wartość e jest dość konserwatywna dla danego gatunku i mieści się w zakresie od 0,01 do 0,03 molCO2 (mol fotonów)-1 Campbell i Norman (1998, s. 237) podają dodatkowe informacje i odniesienia do pełniejszej analizy.
Pomiar f za pomocą ACCUPAR LP-80
Oczywiste jest, że f, ułamek padającego światła przechwytywanego przez koronę roślin, jest krytycznym czynnikiem w określaniu asymilacji. Czynnik ten jest bezpośrednio mierzony za pomocą ACCUPAR LP-80. W środowiskach o ograniczonym dostępie światła można przewidzieć produkcję suchej masy, znając ilość padającego PAR i wydajność konwersji światła, e, a następnie mierząc f w czasie za pomocą LP-80.
Model oparty na wodzie
W sytuacjach ograniczonej ilości wody stosuje się inne równanie. Jest to
Równanie 2
gdzie T to transpiracja, D to deficyt pary wodnej w atmosferze, a k to stała dla danego gatunku i atmosferycznego poziomuCO2. Tanner i Sinclair (1983) oraz Campbell i Norman (1998) podali wyprowadzenia tego równania, ale jego ważność została wielokrotnie potwierdzona w eksperymentach sięgających ponad sto lat wstecz. Przewiduje ono między innymi, że wilgotne regiony będą produkować więcej suchej masy na jednostkę zużytej wody niż obszary suche. Tak więc projekt nawadniania w Wisconsin, powiedzmy, wytworzyłby znacznie więcej suchej masy na jednostkę zużytej wody niż projekt w Arizonie. Chociaż mogą istnieć różnice między poszczególnymi gatunkami w ilości suchej masy produkowanej na jednostkę zużytej wody, każda produkcja suchej masy wymaga znacznej ilości wody. Marzenia o rozkwicie pustyń dzięki inżynierii genetycznej roślin, które wiążą węgiel bez użycia wody, są tylko marzeniami.
Przechwytywanie w modelu opartym na wodzie
Model suchej masy oparty na parowaniu zależy również od przechwytywania światła. Woda utracona przez uprawy obejmuje wodę transpirowaną przez rośliny i wodę odparowaną z gleby. Tylko woda utracona przez transpirację odnosi się do asymilacji węgla. Pomiar T w równaniu 2 zwykle nie jest praktyczny, ale możemy stworzyć prosty model komputerowy, który obliczy go każdego dnia, jeśli znamy deszcz lub nawadnianie oraz niektóre zmienne glebowe i środowiskowe. Na potrzeby modelu musimy zdefiniować wielkość zwaną potencjalną ewapotranspiracją, która jest szybkością utraty wody, gdy zaopatrzenie w wodę nie ogranicza ani parowania, ani transpiracji. Potencjalna transpiracja jest obliczana na podstawie
Równanie 3
Ponieważ parowanie z powierzchni gleby również zużywa wodę, musimy je również obliczyć. Potencjalne parowanie jest obliczane na podstawie
Równanie 4
gdzie Etp to potencjalna ewapotranspiracja. Tak jak poprzednio, f jest ułamkiem promieniowania przechwytywanego przez okap i może być mierzone za pomocą funkcji LP-80. Campbell i Diaz (1988) podają prosty model komputerowy do obliczania Etp, a także algorytmy do obliczania rzeczywistego parowania i transpiracji z potencjalnych wielkości podanych w równaniach 3 i 4.
Wybór odpowiedniego modelu jest łatwy
Najskuteczniejszym sposobem określenia, czy czynnikiem ograniczającym jest światło, czy woda, jest po prostu codzienne uruchamianie obu modeli matematycznych i sprawdzanie, który z nich przewiduje najniższą wartość. Wartość ta jest najlepszym predyktorem produkcji suchej masy w danym dniu, w którym została uruchomiona.
Modelowanie komputerowe BASIC
Modele matematyczne z ograniczeniem światła i wody są trudne do ręcznej obsługi, ale łatwe do zaprogramowania na komputerze. Są one uruchamiane na podstawie łatwo uzyskiwanych danych klimatycznych i mogą być dość dokładnymi predyktorami produkcji suchej masy roślinnej, szczególnie w przypadku upraw jednorocznych. Są one szczególnie przydatne do oceny potencjału produkcyjnego dla poszczególnych środowisk i praktyk kulturowych (Campbell i Diaz, 1988; Kunkel i Campbell, 1987).
Obliczanie przechwytywania frakcji
Frakcja przechwytywania, f używana w obu tych modelach jest wartością uśrednioną dla całych dni. Pomiar dokonywany przez LP-80 jest zazwyczaj dokonywany o określonej porze dnia i nie jest średnią z całego dnia. PodręcznikLP-80 podaje równania i przykład (str. 57) konwersji z pojedynczej obserwacji na średnią dzienną. Strona LP-80 mierzy transmisję promieniowania, biorąc stosunek PAR mierzonego poniżej okapu do PAR mierzonego powyżej. Jest to transmisja przy określonym kącie zenitalnym słońca, ?(θ). Transmisja uśredniona dla całych dni jest taka sama jak transmisja dla promieniowania rozproszonego i wynosi
Równanie 5
gdzie q zależy od leaf area index, rozkładu kąta nachylenia liści i kąta zenitalnego słońca, jak pokazano w instrukcji. Ułamkowe przechwytywanie dla tych modeli wynosi:
Równanie 6
Referencje
Campbell, G. S. i R. Diaz. "Uproszczone modele bilansu wodnego gleby do przewidywania transpiracji upraw". Drought research priorities for the dryland tropics. ICRISAT, Indie (1988): 15-26. Link do artykułu (otwarty dostęp).
Campbell, G. S. i J. M. Norman. An Introduction to Environmental Biophysics (2nd Ed.). New York: Springer, 1998. Link do artykułu.
Kunkel, Robert i Gaylon S. Campbell. "Maksymalny potencjalny plon ziemniaków w Columbia Basin, USA: wartości modelowe i zmierzone". American potato journal 64, nr 7 (1987): 355-366. Link do artykułu.
W jaki sposób obliczanie frakcji wiązki na stronie LP-80 upraszcza pomiary LAI?
Promieniowanie docierające do sondy ACCUPAR LP-80 może pochodzić bezpośrednio z wiązki słonecznej lub być rozproszone z nieba lub chmur. Te dwa źródła mają różny wpływ na architekturę okapu i dlatego muszą być traktowane oddzielnie przy obliczaniu wskaźnika LAI. leaf area index (LAI) z pomiarów transmisji przez koronę. Informacje potrzebne do wykonania obliczeń to frakcja wiązki lub stosunek promieniowania pochodzącego bezpośrednio z wiązki słonecznej do całkowitego promieniowania (wiązka plus rozproszony lub rozproszony PAR) padającego na sondę.
Koniec z cieniowaniem sondy
Poprzednia wersja strony ACCUPAR wymagała od użytkownika pomiaru frakcji wiązki poprzez zacienienie sondy. Strona LP-80 oblicza go przy użyciu dostępnych pomiarów. Zastosowana metoda została zmodyfikowana na podstawie metody opublikowanej przez Spitters et al. (1986) w celu znalezienia frakcji wiązki dla promieniowania całkowitego. Skorelowali oni frakcję wiązki ze stosunkiem zmierzonego całkowitego promieniowania globalnego do promieniowania potencjalnego na poziomej powierzchni poza atmosferą ziemską.
Pomiar PAR powyżej okapu z LP-80 jest całkowitą globalną wartością PAR. Ponieważ szerokość geograficzna i pora dnia są znane, można obliczyć potencjalny PAR (PAR na poziomej powierzchni poza atmosferą ziemską). Stosunek tych dwóch pomiarów jest związany z ułamkiem całkowitego PAR w wiązce słonecznej, tak jak zrobili to Spitters i in. Procedura w LP-80 jest następująca:
Oblicz r, ułamek potencjalnego PAR, który dociera do sondy. Jest to "stała słoneczna" PAR pomnożona przez cosinus kąta zenitalnego, podzielona przez odczyt PAR powyżej okapu. Zakładamy, że "stała słoneczna" PAR wynosi 2550 µmol/m2/s.
Wartość r równa lub wyższa od 0,82 oznacza czyste niebo; wartość równa lub niższa od 0,2 oznacza w pełni rozproszone niebo.
Ułamek r jest wykorzystywany w poniższym wielomianie empirycznym, wyprowadzonym z danych, w celu obliczenia ułamka wiązki:
Makro do wykonania tych obliczeń znajduje się w załączniku poniżej.
LP-80 Teoria oszczędza czas i wysiłek
Podejście to może być mniej dokładne niż bezpośredni pomiar fb, jeśli pomiar ten został wykonany bardzo starannie, ale trudno jest rutynowo wykonywać bezpośrednie pomiary fb, gdy próbuje się zmierzyć przechwytywanie baldachimu lub LAI. W rzeczywistości błędy wprowadzane przez przybliżoną metodę stosowaną w LP-80 są zazwyczaj niewielkie w porównaniu z błędami innych pomiarów. Poniższy wykres przedstawia błąd LAI jako funkcję błędu w szacowaniu frakcji wiązki, przy założeniu, że do wszystkich obliczeń LAI zastosowano stałą frakcję wiązki wynoszącą 0,4.
Błąd ten jest niezależny od LAI. Obliczenia dotyczą kąta zenitalnego 30 stopni. Większe kąty zenitalne mają mniejsze błędy. Wykres pokazuje, że błąd LAI jest zawsze mniejszy niż ± 20%. Dla 10% błędu frakcji wiązki, błąd LAI wynosi około 2%. Trudno jest określić, jak duże mogą być błędy w metodzie LP-80 do obliczania frakcji wiązki, ponieważ zależy to od warunków, ale prawdopodobnie mieszczą się one w zakresie od 10 do 20%. Błąd, który wprowadza to do obliczeń LAI, mieści się zatem w zakresie od 2 do 4%, co jest znacznie mniejsze niż niepewność wynikająca ze zmienności przestrzennej w pomiarze LAI.
Rysunek 1. Błąd LAI jako funkcja błędu w szacowaniu frakcji wiązki, przy założeniu, że do wszystkich obliczeń LAI zastosowano stałą frakcję wiązki wynoszącą 0,4
Odniesienie
Spitters, C. J. T., H. A. J. M. Toussaint i J. Goudriaan. "Oddzielenie rozproszonego i bezpośredniego składnika promieniowania globalnego i jego implikacje dla modelowania fotosyntezy w koronach drzew, część I. składniki promieniowania przychodzącego". Agricultural and Forest Meteorology 38, nr 1-3 (1986): 217-229. Link do artykułu.
Makro Visual BASIC do obliczania frakcji wiązki
Function BeamFraction(Zenith As Single, PAR As Single) As Single
Const pi = 3.14159
Dim r As Single, b As Single
Zenit = Zenit * pi / 180
Jeśli Zenith > 1,5, to
b = 0# 'nighttime
Else
r = PAR / (2550# * Cos(Zenith)) '600 w/m2 * 4.25 umol/w/m2 (.235 MJ/mol)(600 to potencjalny PAR)
Jeśli r > 0,82 Wtedy r = 0,82
If r < 0.2 Then r = 0.2
b = 48,57 + r * (-59,024 + r * 24,835)
b = 1,395 + r * (-14,43 + r * b)
End If
BeamFraction = b
Koniec funkcji
DODATEK B: Dowiedz się więcej o LAI
Dr Steve Garrity omawia Leaf Area Index (LAI). Poruszane tematy obejmują teorię stojącą za pomiarem, metody bezpośrednie i pośrednie, zmienność między tymi metodami, rzeczy do rozważenia przy wyborze metody oraz zastosowania LAI.
Transkrypcja wideo:
Jak obliczyć LAI
W tym wirtualnym seminarium omówimy teorię leaf area index (LAI), różne metody pomiaru LAI i niektóre zastosowania pomiaru LAI. Zaczniemy od zdefiniowania leaf area index. Rysunek 1 przedstawia dwie teoretyczne powierzchnie w lesie lub w uprawie.
Rysunek 1. Schemat koncepcyjny łanu roślinnego, gdzie (a)=1 (b)=3
Działka po lewej stronie to jeden metr z każdej strony lub jeden metr kwadratowy powierzchni ziemi (brązowy kwadrat). Powyżej cały obszar pokryty jest przez liście (zielony kwadrat). Wyobraź sobie naprawdę duży liść pokrywający cały obszar nad poletkiem. Aby obliczyć LAI w lewym przykładzie, wiemy, że powierzchnia gruntu jest równa jednemu metrowi kwadratowemu, a powierzchnia liści jest również równa jednemu metrowi kwadratowemu. LAI jest obliczany jako stosunek powierzchni liści do powierzchni gruntu, w tym przypadku jeden do jednego. Tak więc w tym przykładzie LAI wyniesie jeden.
Po prawej stronie rysunku 1 znajduje się ten sam wykres, ale tym razem z trzema warstwami liści. W tym przypadku mamy jeden metr kwadratowy powierzchni gruntu i trzy metry kwadratowe powierzchni liści, co daje nam stosunek powierzchni liści do powierzchni gruntu wynoszący trzy do jednego. Tak więc w tym przypadku LAI wynosi trzy.
Dlaczego warto mierzyć LAI?
LAI nie jest skomplikowaną koncepcją do zrozumienia i chciałbym omówić, dlaczego mierzymy leaf area index lub dlaczego jest to przydatne. LAI jest jedną z tych zmiennych, które są dość wszechobecne, co oznacza, że są używane wszędzie. Dzieje się tak dlatego, że jest prosta, ale także niezwykle opisowa.
To jest mapa globalnego LAI uzyskana z danych satelitarnych (patrz kod czasowy webinaru: 2:16). Obszary o wysokim LAI są reprezentowane przez kolor ciemnozielony, a obszary o niskim LAI są jasnozielone. Zauważ, że w tropikach wokół równika znajdują się jedne z najgęstszych lasów o najwyższym LAI na Ziemi. Z kolei na północ lub południe od równika, gdzie występuje wiele naszych pustyń, LAI jest bardzo niski. Następnie, przesuwając się dalej na północ lub południe w strefach umiarkowanych (strefy borealne), LAI ponownie wzrasta. Wzorce LAI na tej mapie odzwierciedlają wiele procesów i wiele zmiennych. Dostępność wody lub światła może wyjaśniać niektóre z tych wzorców, ale na tym jednym przykładzie widać, że LAI bardzo dobrze opisuje wzorce roślinności na świecie.
Oto kilka innych powodów, dla których LAI jest tak ważny:
Zbieranie światła z okapu (produktywność, akumulacja biomasy, wzrost upraw)
Fenologia
Struktura czaszy
Transpiracja
Skalowanie procesów i nie tylko
LAI jest związany ze zbieraniem światła. Im więcej liści w łanie, tym większa zdolność do pochłaniania energii świetlnej ze słońca. Ta energia świetlna jest następnie wykorzystywana do napędzania produktywności roślin (produktywności pierwotnej) poprzez pobieranie i przekształcanie dwutlenku węgla z atmosfery w węglowodany. Jest to związane z akumulacją biomasy oraz wzrostem upraw i lasów.
LAI jest również używany jako wskaźnik fenologii, gdzie fenologia opisuje po prostu cykl życia roślin. Na przykład w lasach liściastych każdego roku liście wypuszczają pędy, rosną, rozwijają się, dojrzewają i w końcu obumierają. Wszystkie te procesy można opisać poprzez śledzenie leaf area index w czasie.
LAI jest również powszechnie stosowany jako miara struktury korony lub sposób na odróżnienie struktury jednej korony od drugiej. Jest on przydatny w dwóch powiązanych parametrach: transpiracji i procesach skalowania.
Rysunek 2. Procesy wymiany zachodzą na powierzchni liścia
Weźmy na przykład liść (rysunek 2). W liściu tym zachodzi wiele procesów fizjologicznych, które oddziałują z otaczającą atmosferą na powierzchni liścia. Interakcje te polegają na wymianie zarówno masy, jak i energii. Jeśli rozumiemy te procesy wymiany na poziomie liścia i wiemy, ile liści znajduje się w czaszy poprzez LAI, daje nam to wygodną metodę skalowania tych procesów do poziomu czaszy i dalej.
Jak mierzyć LAI
Istnieją dwa główne podziały metod pomiaru LAI: metody bezpośrednie i metody pośrednie. Bezpośrednie metody pomiaru LAI zazwyczaj wiążą się z destrukcyjnym wycinaniem koron drzew: ścinaniem drzew lub przycinaniem biomasy. Jednym ze sposobów, który nie jest tak destrukcyjny, jest użycie pułapek na ściółkę do wychwytywania liści, które starzeją się i opadają z roślin. Z kolei metody pośrednie nie mierzą LAI bezpośrednio, ale mierzą inne powiązane zmienne. Powiązane zmienne są następnie wykorzystywane jako zamienniki LAI lub do bezpośredniego modelowania LAI. Metody pośrednie, które omówię podczas tego seminarium, to fotografia półkulista, inwersja PAR (która wykorzystuje pomiary promieniowania przepuszczanego przez koronę drzew) i odbicie spektralne (podejście odgórne wykorzystujące czujniki nad koroną drzew).
LAI: metody bezpośrednie
Jak wspomniano, niszczące zbiory są powszechne w bezpośrednich metodach LAI. W lesie wiąże się to z wycinaniem drzew i usuwaniem całego materiału liściowego z tych drzew: pracochłonny, żmudny proces, który usuwa również znaczną ilość materiału z okapu.
Rysunek 3. Bezpośrednia metoda niszcząca, w której badacze zbierają cały materiał liściowy z poletka
Rysunek 3 ilustruje bardzo krótki baldachim, w którym badacze wyznaczają okrągłe poletko na ziemi i zbierają cały materiał liściowy z tego poletka. W tym przypadku zastosowanie metody destrukcyjnej może być jedynym sposobem pomiaru LAI tylko dlatego, że baldachim jest tak krótki.
Innym sposobem bezpośredniego pomiaru LAI jest użycie pułapek na ściółkę. W lasach liściastych każdej jesieni liście starzeją się i opadają na ziemię. Pułapki na ściółkę można umieścić wokół okapu, aby uchwycić niektóre z tych liści. Badacze mogą następnie okresowo pobierać próbki liści (tj. wyciągać je z pułapki i zabierać do laboratorium w celu analizy).
Zarówno w przypadku niszczących metod zbioru, jak i pułapek na ściółkę, po wyodrębnieniu materiału liściowego z rośliny należy zmierzyć ilość zebranego obszaru liści. Jedną z popularnych metod jest Licor Li 3100, który jest zasadniczo skanerem optycznym. Badacz przepuszcza każdy liść przez skaner i mierzy jego powierzchnię. Po zeskanowaniu wszystkich liści badacz może zsumować powierzchnię i podzielić ją przez powierzchnię gruntu, aby uzyskać miarę LAI. Jedną z unikalnych zalet tej metody jest to, że pozwala ona na pomiar specyficzny dla danego gatunku leaf area index. Jest to pomocne w systemach niezarządzanych lub zadaszeniach mieszanych gatunków w celu zrozumienia wkładu każdego gatunku w całkowity LAI zadaszenia. Badacz może zebrać gatunki A, B i C, a następnie przeanalizować ich powierzchnię liści niezależnie za pomocą skanera.
LAI: metody pośrednie
Wszystkie pośrednie metody LAI omówione w tym webinarium opierają się w pewien sposób na pomiarze interakcji światła z baldachimem, więc najpierw krótki przegląd tego, jak światło może oddziaływać z baldachimem. Istnieją trzy losy światła w baldachimie.
Transmisja: Światło słoneczne jest przepuszczane przez cały baldachim.
Absorpcja: Światło słoneczne pochłaniane lub przechwytywane przez baldachim, a energia jest wykorzystywana w procesie fotosyntezy.
Odbicie: Światło słoneczne uderza w górną część baldachimu i jest odbijane z powrotem do atmosfery i w przestrzeń kosmiczną.
Możemy zmierzyć dwie z tych wielkości: transmitancję i współczynnik odbicia. Absorbancja jest niemierzalna, ponieważ energia jest wykorzystywana przez roślinę.
Fotografia półkulista
Fotografia hemisferyczna to metoda wykorzystująca pomiar przepuszczanego światła do oszacowania LAI. Jest to metoda, która istnieje już od dłuższego czasu i ma ugruntowaną pozycję. Polega ona na użyciu aparatu z obiektywem typu rybie oko, przymocowaniu całego aparatu do platformy poziomującej, a następnie skierowaniu go w górę, tak aby znajdował się pod baldachimem skierowanym w stronę nieba.
Rysunek 4. Fotografia półsferyczna z mieszanego lasu liściastego przy użyciu obiektywu typu rybie oko w aparacie cyfrowym
Kamera rejestruje obraz łanu od dołu w półkuli takiej jak na rysunku 4. Widać więc, że siedem obrazów wzdłuż dolnej krawędzi (patrz obrazy w kodzie czasowym 13:08 w webinarium) to sekwencja czasowa zdjęć, które zostały zebrane z tego samego miejsca w koronie lasu liściastego od bardzo wczesnej wiosny do mniej więcej połowy lata. Wizualnie zdjęcia te pokazują, że wczesną wiosną w koronach drzew jest niewiele materiału liściowego lub nie ma go wcale. W połowie lata liście są już w pełni rozwinięte i dojrzałe.
Fotografia półsferyczna jest wyjątkowa, w przeciwieństwie do niektórych innych metod, o których będę mówić, ponieważ obraz czaszy jest niezwykle bogatym zestawem danych. Dzieje się tak, ponieważ zawiera zarówno komponent przestrzenny, jak i kolorystyczny. Zapewnia również archiwum lub zapis danych, które można następnie ponownie przeanalizować (tj. możliwe jest użycie innej metody do analizy obrazów w miarę zmian teorii i technologii). Podczas gdy w przypadku innych metod mierzysz pewną wartość i nie możesz jej ponownie zmierzyć.
Inną zaletą fotografii półkulistej jest to, że oprócz LAI można również zmierzyć zestaw innych zmiennych związanych ze strukturą korony. Na przykład, wykreśliłem tutaj hipotetyczną ścieżkę słoneczną: pozycję lub ścieżkę, którą słońce pokonuje na niebie w danym dniu. Można wykorzystać te informacje, aby wykreślić, gdzie będzie znajdować się słońce, a następnie oszacować, kiedy w przykładowej lokalizacji może wystąpić plama słoneczna i jaki może być czas jej trwania. Może to być ważne, jeśli chcesz zbadać, w jaki sposób LAI jest powiązany z przepuszczalnością światła i jak wpływa to na dostępność światła dla gatunków podszytu. Naukowcy wymyślili wiele innych sposobów wydobywania informacji ze zdjęć półkulistych niż tylko leaf area index.
Aby przeanalizować zdjęcia półkuliste, surowe zdjęcie jest przetwarzane za pomocą oprogramowania w celu uzyskania szacunkowej wartości LAi lub innej zmiennej. Odbywa się to za pomocą progowania. Ideą progowania jest rozróżnienie pikseli zajmowanych przez liście od pikseli zajmowanych przez niebo. Zauważ, że w lewym górnym rogu znajduje się nieprzetworzony obraz (patrz kod czasowy webinaru 15:14). Pozostałe siedem obrazów przedstawia różne wartości progowe, które zostały zastosowane do tego obrazu. Jest to moim zdaniem pięta achillesowa fotografii półkulistej, ponieważ różni obserwatorzy mogą wybierać różne wartości progowe w oparciu o to, co mówi im oko. Ponadto różne zautomatyzowane metody wykrywania progu mogą dawać różne wyniki. Analiza zdjęć półkulistych jest więc dość subiektywna, co może utrudniać porównywanie zdjęć wykonanych w różnym czasie lub gdy w przetwarzanie danych zaangażowane są różne osoby.
Podczas fotografowania półkulistego należy unikać robienia zdjęć, gdy tarcza słoneczna prześwituje przez baldachim. Wynika to z faktu, że tuż wokół tarczy słonecznej będzie bardzo jasna plama, a jeśli spróbujesz ocenić różnicę między jasnym tłem, jasnym niebem i baldachimem, nie docenisz, ile baldachimu jest tam z powodu tej jasnej plamy. Ponadto, ponieważ obraz jest zbierany, gdy słońce świeci bezpośrednio na baldachim, w baldachimie będą rzucane cienie, co bardzo utrudni rozróżnienie progu jasności związanego z niebem i baldachimem. Wreszcie, jeśli na zdjęciu występują zmienne chmury, obszary zachmurzone będą bardzo jasne, podczas gdy tło nieba będzie nieco ciemniejsze. To sprawia, że bardzo trudno jest wybrać próg, który odróżnia baldachim od braku baldachimu. Z tych wszystkich powodów zaleca się wykonywanie zdjęć półkulistych tylko w warunkach równomiernego rozproszenia światła lub równomiernego zachmurzenia. Inną porą dnia, która jest odpowiednia, jest bardzo wcześnie lub bardzo późno, gdy słońce jest nisko lub poniżej horyzontu, aby wyeliminować problemy z zanieczyszczeniem obrazu przez tarczę słoneczną.
Jakie zastosowania są więc odpowiednie dla fotografii półkulistej? Pole pszenicy prawdopodobnie nie jest dobrym miejscem do robienia zdjęć półkulistych, ponieważ łan pszenicy jest dość nisko rosnący i trudno byłoby umieścić aparat, obiektyw, platformę poziomującą i statyw w całości pod łanem. Fotografia półsferyczna dobrze sprawdza się w wysokich baldachimach, takich jak baldachim leśny, ponieważ łatwo jest zmieścić sprzęt pod wszystkimi liśćmi w baldachimie.
LP-80Światło przechodzące i prawo Beera
Z koncepcyjnego punktu widzenia można stwierdzić, czy znajdujemy się w rzadkim okapie, ponieważ jest tam bardzo mało liści, a w podszycie rzadkiego okapu jest zwykle znacznie jaśniej. Natomiast w przypadku bardzo gęstego okapu można oczekiwać, że duża część światła zostanie pochłonięta lub odbita i nie zostanie przekazana do podszytu.
Rysunek 5. Istnieje zależność między przepuszczalnością światła a powierzchnią liści
Korzystając z tych podstawowych obserwacji, można zauważyć, że istnieje pewien związek między przepuszczalnością światła a powierzchnią liści. Jest to sformalizowane przez prawo Beera, a dla celów LAI należy rozważyć formę prawa Beera dotyczącą energii świetlnej w postaci promieniowania fotosyntetycznie czynnego lub PAR.
Równanie 1
PARt jest transmitowanym paskiem, który może być mierzony w dolnej części czaszy. Będzie to funkcja padającego PAR(PARi) lub ilości promieniowania fotosyntetycznie czynnego padającego na górną część okapu. Dwa kolejne parametry to k i z, gdzie k to współczynnik ekstynkcji, a z to połączenie ścieżki przez medium tłumiące. W tym przypadku ośrodkiem tłumiącym byłby sam baldachim. Tak więc prawo Beera w tej formie jest podstawą sposobu, w jaki wykorzystujemy pomiary przepuszczanego światła do oszacowania LAI. W szczególności zilustruję model matematyczny używany przez METER Accupar LP-80 (równania 2 i 3).
Równanie 2
W równaniu 2 w lewym górnym rogu L to leaf area index, a pierwszym parametrem, którym chciałbym się zająć, jest obliczenie k, czyli współczynnika ekstynkcji w modelu. W prawym dolnym rogu równania 2 znajduje się podmodel z dwoma parametrami: chi (X) i theta(𝚹). Theta to po prostu kąt zenitalny Słońca w momencie wykonywania pomiaru.
Rysunek 6. Kąty zenitalne słońca zmieniają się w ciągu dnia. Obserwator jest zwrócony w stronę równika.
W ciągu dnia zmienia się kąt zenitalny słońca. Na rysunku 6 słońce znajduje się w różnych miejscach na niebie. Wczesnym rankiem (po lewej) słońce znajduje się niżej na niebie w porównaniu z okresami bliżej południa. To samo dzieje się pod koniec dnia. Theta jest ważna dla opisania długości ścieżki promieniowania wiązki (droga fotonów bezpośrednio od słońca do obserwatora do jakiegoś punktu w czaszy).
Zauważ, że na początku lub pod koniec dnia długość ścieżki jest znacznie dłuższa niż w środku dnia. Tak więc kąt zenitalny słońca jest obliczany po prostu na podstawie pory dnia i znajomości położenia geograficznego. Na stronie LP-80 parametry te są obliczane automatycznie na podstawie wprowadzonych przez użytkownika wartości czasu i lokalizacji, dlatego podczas konfigurowania strony LP-80 bardzo ważne jest prawidłowe wprowadzenie obu tych wartości.
Równanie 3
Kolejną zmienną w modelu współczynnika ekstynkcji (równanie 2 w prawym dolnym rogu) jest chi (XWartość chi opisuje rozkład kąta nachylenia liści w czaszy. Każdy baldachim jest mieszanką liści o orientacji poziomej, pionowej lub gdzieś pomiędzy poziomą a pionową. Rysunek 7 to wykres przedstawiający rozkład kątów liści w trzech różnych baldachimach.
Rysunek 7. Rozkład kątów liści w trzech różnych baldachimach (Campbell i Newman, 1998)
Należy zauważyć, że wartości chi w pionowych baldachimach są niższe niż jeden. Im bardziej pionowy jest rozkład kąta nachylenia liści, tym bardziej chi zbliża się do zera. W poziomych zadaszeniach chi zbliża się do nieskończoności. Zazwyczaj w tym przypadku wartości chi są większe niż jeden (tj. wartości od jednego do pięciu są powszechne w poziomych zadaszeniach). Daszki sferyczne to daszki z mieszanką rozkładów pionowych i poziomych. Są to najczęściej spotykane rozkłady kątów liści występujące w przyrodzie. Mają one wartości chi bliskie lub równe jeden. Program LP-80 domyślnie używa wartości chi równej jeden. Można to zmienić, ale w większości przypadków można uniknąć domyślnego ustawienia.
Rysunek 8. Wpływ wartości chi lub rozkładu kąta nachylenia liści na współczynnik ekstynkcji w zależności od kąta zenitalnego słońca (Campbell i Norman, 1998)
Wykres w lewym dolnym rogu rysunku 8 pokazuje, jak wartość chi lub rozkład kąta nachylenia liści wpływa na współczynnik ekstynkcji zależny od kąta zenitalnego słońca. Na przykład zauważ, że przy chi równym zero (całkowicie pionowy baldachim) i słońcu bezpośrednio nad głową (kąt zenitalny wiązki jest równy zero), współczynnik ekstynkcji jest równy zero, co oznacza, że całe promieniowanie przechodzi przez baldachim. Żadne z nich nie jest pochłaniane ani odbijane. Jest ono w 100% transmitowane.
Porównajmy to z przypadkiem, w którym wszystkie listki są idealnie poziome (chi równa się nieskończoności). Wtedy współczynnik ekstynkcji nie zależy od kąta zenitalnego wiązki. Ma to sens, jeśli pomyślimy o idealnie poziomym liściu. Nie ma znaczenia, pod jakim kątem pada na niego promieniowanie słoneczne. Współczynnik ekstynkcji będzie niezmienny.
Wykres w prawym dolnym rogu rysunku 8 przedstawia transmisję w zależności od kąta zenitalnego słońca. Należy zauważyć, że pozioma transmisja baldachimu jest taka sama bez względu na kąt zenitalny. Na drugim biegunie, dla pionowych baldachimów, transmisja jest równa jeden, gdy słońce znajduje się bezpośrednio nad głową, i jest pełna, gdy mamy bardzo niski kąt padania promieni słonecznych (słońce na horyzoncie). Ma to sens, gdy pomyślimy o pionowym liściu i słońcu bezpośrednio nad głową. Liść nie rzuca cienia, natomiast jeśli słońce pada z boku, następuje całkowita absorpcja i brak transmisji promieniowania.
Czego możemy się z tego dowiedzieć? W lewym górnym rogu rysunku 8 znajdują się 3 różne zadaszenia z bardzo różnymi rozkładami kąta nachylenia liści, a tym samym różnymi wartościami chi w zakresie od 0,5 do trzech. Jeśli jednak spojrzeć na oba rysunki w lewym i prawym dolnym rogu, nie ma dużej różnicy ani we współczynniku ekstynkcji, ani w transmisji między tymi wartościami chi. Tak więc model leaf area index nie jest bardzo wrażliwy na wartość chi, zwłaszcza wartości chi w zakresie od 0,5 do 2.
Tak więc błędne oszacowanie chi może być źródłem błędu, ale tylko w skrajnych przypadkach - jeśli mamy do czynienia z baldachimem, który jest wyjątkowo poziomy lub bardzo pionowy. Jeśli nie mamy do czynienia z żadną z tych skrajności, wartość chi w okolicach jednego będzie odpowiednia do oszacowania LAI.
Równanie 4
Wracając do modelu LAI (równanie 4), Fb to frakcja wiązki i jest obliczana jako stosunek między rozproszonym PAR (promieniowaniem fotosyntetycznie czynnym) a bezpośrednim PAR.
Rysunek 9. Rozproszony PAR vs. bezpośredni PAR
Rysunek 9 ilustruje, co to oznacza. Po lewej stronie znajdują się typowe warunki bezchmurnego nieba z białymi liniami reprezentującymi promieniowanie rozproszone (promieniowanie rozproszone w atmosferze przez aerozole w innych cząstkach). Promieniowanie to jest rozpraszane do pewnego miejsca w czaszy, gdzie możemy mierzyć światło przechodzące. Zauważ również promieniowanie wiązki (promieniowanie pochodzące bezpośrednio od słońca) na tym samym zdjęciu, które dominuje w warunkach czystego nieba. Widzimy więc, że po lewej stronie Fb byłoby bardzo niskie, ponieważ dominuje bezpośredni składnik PAR.
Porównaj to z obrazem po prawej stronie, gdzie w atmosferze znajdują się chmury lub ciężkie aerozole. Występuje tam większe rozproszenie, a mniej promieniowania wiązki przenika przez chmury do miejsca obserwacji poniżej czaszy. W tym przypadku Fb byłoby znacznie wyższe (zbliżając się do 1), ponieważ całkowicie eliminujemy składnik promieniowania wiązki.
Co to oznacza? Fb jest ważny, ponieważ opisuje rozkład kątów przenikania fotonów do czaszy. Fb współdziała z rozkładem kąta nachylenia liści, aby opisać prawdopodobieństwo, że foton przeniknie lub zostanie przepuszczony przez cały baldachim. Na przykład, w bardzo słoneczny dzień można zaobserwować wiele ostrych cieni. Rzucane cienie są niezwykle głębokie i ciemne. Podczas gdy w pochmurny dzień trudniej jest znaleźć silne cienie. Wynika to z bardziej równomiernego rozkładu kątów promieniowania padającego na obiekty, które mogą rzucać cień. Podobnie jest z liśćmi w baldachimie.
Rysunek 10. Tau to stosunek transmitowanego i padającego PAR
Kolejnym terminem, który omówimy, jest tau(𝛕), czyli stosunek promieniowania fotosyntetycznie czynnego transmitowanego do padającego. Wartość tau jest prawdopodobnie najważniejszym składnikiem modelu LAI. Model LAI jest najbardziej wrażliwy na tau. Jest to składnik, który stanowi rdzeń pomiaru podczas korzystania z tego modelu. Na rysunku 10 mierzymy promieniowanie padające w górnej części baldachimu za pomocą PAR sensor. Następnie poniżej baldachimu używamy LP-80 do pomiaru ilości światła przepuszczanego przez baldachim. Model ten wymaga pomiarów zarówno powyżej, jak i poniżej okapu.
Jeśli baldachim jest bardzo wysoki, należy znaleźć polanę lub dużą lukę, w której można umieścić urządzenie PAR sensor i użyć go do pomiaru promieniowania padającego. Możesz albo umieścić PAR sensor na polanie, która jest stale rejestrowana, albo zabrać sam LP-80 na polanę, uzyskać odczyt padającego promieniowania, a następnie zabrać go z powrotem do czaszy, aby zmierzyć promieniowanie transmitowane.
Jeśli pracujesz w warunkach częściowego zachmurzenia lub warunki na niebie szybko się zmieniają, wówczas chcesz dość często aktualizować odczyt promieniowania padającego: w zasadzie za każdym razem, gdy zmieniają się warunki na niebie (a tym samym poziom oświetlenia otoczenia). Z tego powodu, jeśli obawiasz się wahań poziomu oświetlenia otoczenia, zalecam niezależne rejestrowanie zarówno promieniowania padającego, jak i transmitowanego jednocześnie, aby zawsze uwzględniać zmiany poziomu oświetlenia otoczenia i nie wprowadzać żadnego źródła błędu do obliczeń LAI.
Strona LP-80 doskonale nadaje się do punktowego lub okresowego pobierania próbek. W celu ciągłego monitorowania zmian LAI, innym podejściem byłoby użycie czujników PAR zarówno powyżej, jak i poniżej okapu. Czujniki PAR poniżej okapu zasadniczo zastępują LP-80 na rysunku 10. Różnica polega na tym, że czujniki PAR mogą stale rejestrować, co zapewnia ciągły pomiar transmitowanego promieniowania do wprowadzenia do modelu LAI.
Rysunek 11. A to absorpcja PAR liścia (wykres: www.photobiology.info)
Ostatnim terminem w modelu LP80 leaf area index (LAI) jest A, który jest absorpcją liści w obszarze PAR (fotosyntetycznie aktywnym) widma elektromagnetycznego.
W LP-80, A jest ustalone na wartość 0,9, co jest bardzo dobrym oszacowaniem absorbancji. W przypadku większości zadaszeń absorbancja nie ulega dużym zmianom. W niektórych ekstremalnych przykładach może być inaczej. Na przykład, jeśli liście są bardzo młode, ich absorbancja może być znacznie niższa niż 0,9. A gdy są starzejące się, może być niższa niż 0,9. Z pewnością w przypadku bardzo owłosionych lub wyjątkowo woskowatych liści wartość absorbancji może być nieco niższa niż 0,9. Jednak poza skrajnymi przypadkami, wartość 0,9 jest bardzo dobrym oszacowaniem absorbancji liści. Wartości nieznacznie odbiegające od 0,9 nie będą miały dramatycznego wpływu na obliczenia LAI.
Odbicie: pośrednia metoda obliczania LAI
W przypadkach, gdy LAI jest bardzo niski, zazwyczaj występuje równomierny współczynnik odbicia zarówno w widzialnej, jak i bliskiej podczerwieni części widma. Wraz ze wzrostem LAI zmniejsza się współczynnik odbicia światła widzialnego, podczas gdy współczynnik odbicia w bliskiej podczerwieni ma tendencję do wzrostu. Istnieje więc związek między współczynnikiem odbicia światła widzialnego i bliskiej podczerwieni a LAI, który możemy wykorzystać do oszacowania LAI.
Rysunek 12. Dane dotyczące współczynnika odbicia
Na rysunku 12 należy zauważyć, że współczynnik odbicia zależy od długości fali. Wykres w lewym dolnym rogu obejmuje zarówno zakres widzialny (od 400 do 700 nanometrów), jak i część zakresu bliskiej podczerwieni (powyżej 700 nanometrów) widma elektromagnetycznego. Widać, że widmo jest zbierane dla tego samego baldachimu, ale przy różnych wartościach leaf area index (LAI). To, co opisuję, to spadek współczynnika odbicia światła widzialnego wraz ze wzrostem LAI i wzrost współczynnika odbicia w bliskiej podczerwieni wraz ze wzrostem LAI.
Istnieją indeksy wegetacji lub kombinacje różnych pasm, które pozwalają nam oszacować różne biofizyczne zmienne baldachimu. Jednym z powszechnych indeksów jest znormalizowany indeks różnicowy wegetacji (NDVI).
Rysunek 13. Każdy baldachim ma unikalną relację NDVI-LAI
NDVI jest formułowany przy użyciu odbitych wartości promieniowania czerwonego i bliskiej podczerwieni, a wykazano, że NDVI jest powiązany z leaf area index. Rysunek 13 przedstawia czujnik odbicia spektralnego na szczycie czaszy, który stale monitoruje odbite promieniowanie w dwóch pasmach. Dwa porty mierzą promieniowanie czerwone i bliskiej podczerwieni. Jeśli jednak chcemy użyć tej wartości NDVI jako bezpośredniego oszacowania LAI lub jako sposobu oszacowania bezwzględnej wartości LAI, musimy opracować związek z jakąś niezależną miarą LAI.
Na przykład, moglibyśmy użyć LP-80 do obliczenia LAI na podstawie pomiarów transmitowanego promieniowania, a następnie kolokować czujnik odbicia spektralnego zbierający wartości NDVI . Jeśli zbierzemy wystarczającą liczbę tych wartości w czasie lub w przestrzeni, możemy opracować zależność regresji liniowej (Rysunek 13, u góry po lewej). Następnie moglibyśmy wykorzystać kolejne wartości NDVI w tym równaniu empirycznym do obliczenia leaf area index bez konieczności korzystania z niezależnego źródła (LP-80) LAI dla wszystkich kolejnych pomiarów.
Być może nie potrzebujesz bezwzględnych wartości LAI i masz inny powód do pomiaru LAI. Rysunek 14 przedstawia kilka przykładów wykorzystania strony NDVI jako wskaźnika zastępczego LAI bez konieczności podawania bezwzględnej wartości LAI.
Rysunek 14. NDVI może być używany jako wskaźnik zastępczy dla LAI lub dla powiązanych zmiennych (Ryu et al. (2010) Ag for Met).
W tym przypadku badacz mierzył zarówno fotosyntezę NDVI , jak i fotosyntezę w koronach drzew na użytkach zielonych przez cały rok. W lewym górnym panelu wartości NDVI są wykreślone na zielono, a następnie fotosynteza jest pokazana za pomocą otwartych okręgów. Widać, że trajektoria czasowa fotosyntezy jest bardzo dobrze śledzona przez NDVI. Pokazuje on, jak można opracować równanie regresji, które wiąże wartości NDVI z fotosyntezą w koronach drzew. W tym przypadku, leaf area index jest jednym z silnych czynników wpływających na fotosyntezę na tych jednorocznych użytkach zielonych. Ale zamiast próbować modelować fotosyntezę w koronach drzew na podstawie LAI, po prostu używa NDVI jako wskaźnika zastępczego.
Podobnie moglibyśmy rozważyć aplikację fenologiczną. Wykresy w prawym dolnym rogu rysunku 14 przedstawiają niektóre dane zebrane z lasu liściastego przez siedem lat z LAI i NDVI mierzonymi w różnych odstępach czasu. Subiektywnie widzimy, że NDVI bardzo dokładnie śledzi czasową dynamikę leaf area index . W tym przypadku moglibyśmy więc zastąpić miarę LAI wskaźnikiem NDVI.
Rozważania dotyczące LAI: pobieranie próbek i skalowanie
Nie myśl, że możesz zmierzyć LAI w jednym miejscu i uzyskać jedną wartość reprezentatywną dla całego baldachimu. To nie działa w ten sposób. Jednym z założeń, które zwykle przyjmujemy w modelu typu LAI, jest to, że liście są losowo rozmieszczone w łanie. Prawie nigdy tak nie jest. Zawsze istnieje pewien stopień zbrylenia, który występuje tylko ze względu na wzór rozgałęzień i sposób, w jaki liście, gałęzie i drzewa są rozmieszczone w okapie.
Jednym z najprostszych sposobów na obejście negatywnych skutków zbrylania lub zmienności przestrzennej jest zwiększenie wielkości próby.
Rysunek 15. Artystyczne odtworzenie lotniczego obrazu pola (Colombo et al. (2003) Rem. Sens. Env)
Po lewej stronie rysunku 15 znajduje się artystyczne odtworzenie zdjęcia lotniczego kilku różnych pól uprawnych. Po prawej stronie znajduje się odtworzenie obrazu z systemu obrazowania używanego do zbierania danych NDVI z tego samego obrazu pola uprawnego, a następnie przekształcenia go w dane NDVI , a następnie w leaf area index. Widać, że na tym obrazie występuje szeroki zakres wartości LAI w różnych jednostkach zarządzania. Obrazowanie daje nam poczucie niejednorodności przestrzennej, ale metody, o których mówiliśmy, są bardziej dyskretne pod względem obszaru, który reprezentują. Możemy temu zaradzić, zbierając wiele próbek w naszym obszarze badań, aby spróbować uchwycić zmienność przestrzenną. Następnie można przyjąć pewnego rodzaju średnią przestrzenną, aby przedstawić LAI na całym obszarze.
A może po prostu interesuje nas zrozumienie, jaka jest zmienność LAI na całym obszarze. Obraz, który pokazałem na początku tego seminarium (Rysunek 1) na temat globalnego rozkładu leaf area index został uzyskany z danych satelitarnych. Ale jak możemy zaufać tym wartościom? Musimy mieć jakiś sposób na sprawdzenie tych wartości. W jaki sposób? Można zainstalować czujnik NDVI nad baldachimem, wykonujący bardzo szczegółowe pomiary na poziomie lokalnym. Sprawdzić go z tym, co dają nam dane satelitarne, a następnie przypisać pewien poziom zaufania do tego, co widzimy poza naszym obszarem próbkowania, korzystając z danych satelitarnych.
Rysunek 16. LAI w koronach drzew lasów liściastych wiosną (Garrity et al. (2008) ESA)
Należy pamiętać, że nie wszystkie metody dadzą takie same wyniki. Rysunek 16 przedstawia niektóre dane, które zebrałem kilka lat temu wiosną w lesie liściastym. Wykorzystałem cztery różne metody: fotografię półkulistą, LAI 2000 i czujnik kwantowy (PAR sensor). Następnie użyłem satelity MODIS (dostarczają produkt LAI) i zestawiłem go z niektórymi z moich pomiarów i porównałem wszystkie cztery. Należy zauważyć, że w danym dniu istnieje dość duża zmienność między szacunkami dostarczonymi przez dowolną z tych metod. Może to więc stanowić wyzwanie podczas porównywania jednej metody z drugą. Niektóre metody mają tendencję do lepszego porównywania się ze sobą. Na przykład, nie miałem LP-80 dla tego badania, ale obecnie opublikowano około trzech lub czterech różnych artykułów, które pokazują, że LAI 2000 i LP-80 zazwyczaj dają wartości, które są do siebie bardzo podobne. Teoretycznie czujniki kwantowe powinny być bardzo zbliżone zarówno do LP-80 , jak i LAI 2000.
Prawda jest taka, że żadna z tych metod nie podaje prawidłowej wartości bezwzględnej. W tym przypadku użyliśmy pułapek na ściółkę, które były najbardziej bezpośrednim sposobem oszacowania rzeczywistego LAI. W tym przypadku wynosił on nieco poniżej 4,0. Widać więc, że przynajmniej w okresie dojrzałości żadna z tych metod nie uzyskała dokładnej wartości. Należy więc zachować ostrożność przy porównywaniu różnych metod lub po prostu zrozumieć, że istnieje zmienność w zależności od metody.
Jedno ze źródeł zmienności, którego możemy uniknąć, zostało przedstawione na tym obrazku (patrz kod czasowy webinaru 46.01). Obraz ten demonstruje kilka koncepcji, o których rozmawialiśmy. Strumienie światła przenikają przez baldachim, kontrastując z niektórymi zacienionymi obszarami. Widać, że cała ta dynamika światła jest kontrolowana przez ilość materiału liściowego w baldachimie i jego rozmieszczenie w baldachimie. Jeśli więc mamy pojedynczy PAR sensor , który mierzy transmitowane promieniowanie w tym baldachimie. Jeśli umieścimy go po prawej stronie, w tym momencie, w którym wykonano zdjęcie, odczytamy bardzo wysokie wartości przepuszczanego światła. Ale jeśli mamy inny PAR sensor tutaj w cieniu, zobaczymy bardzo niskie wartości przepuszczanego światła. Musimy więc zwracać uwagę na zmienność przestrzenną mierzonego okapu.
Rysunek 17. Dane PAR (Garrity, et al. (2011) Ag For Met)
Rysunek 17 pokazuje niektóre dane, które pokazują, jak wyglądają, gdy patrzymy na poszczególne czujniki PAR. W tym przypadku było ponad 30 czujników PAR rozmieszczonych pod okapem lasu liściastego. Z biegiem czasu wszystkie mają tendencję do śledzenia siebie nawzajem, ale bezwzględna wartość transmitowanego promieniowania może być bardzo różna w zależności od lokalizacji. Jeśli więc używamy światła przechodzącego jako oszacowania LAI, jakiego śladu używamy do oszacowania LAI? Odpowiedź zależy od tego, jaki jest nasz cel. Jeśli próbujemy po prostu uzyskać średnie poczucie tego, czym jest LAI, być może weźmiemy średnią przestrzenną wszystkich tych wartości.
Inną rzeczą, na którą należy zwrócić uwagę, jest to, że te czynniki zbrylania i zmienności przestrzennej są prawdziwymi źródłami błędów. Jednak LP-80 uwzględnia to w sposobie pozyskiwania pomiarów światła przechodzącego. Posiada on różdżkę, która wychodzi z urządzenia ręcznego, a ta różdżka ma około 80 centymetrów długości i 80 niezależnych czujników PAR w tej różdżce. Tak więc odczyty LP-80 są średnią przestrzenną wszystkich czujników w różdżce.
Rysunek 18. Strona ACCUPAR LP-80 mierzy PAR i LAI
Kilka lat temu niektórzy badacze wykazali, że w koronach drzew, w których występują skupiska, jeśli weźmie się średnią z liniowego transektu, ma się tendencję do zmniejszania ilości błędów związanych ze skupiskami, a strategia ta jest już fizycznie wbudowana w LP-80.
Jeśli korzystasz z PAR sensor, jednym z podejść jest upewnienie się, że zbierasz wystarczającą liczbę próbek, które reprezentują przestrzenną niejednorodność przepuszczalności światła, co oczywiście jest związane z LAI.
Dlaczego mierzysz LAI?
Przed dokonaniem pomiaru należy zastanowić się, dlaczego mierzy się LAI. Czy naprawdę interesuje Cię leaf area index? A może interesuje Cię jakaś powiązana zmienna? Na przykład, niektórzy badacze szacują LAI, aby móc częściej szacować przepuszczane lub pochłaniane światło, ponieważ próbują oszacować produktywność korony lub fotosyntezę. Pytanie brzmi, po co szacować LAI, aby oszacować absorpcję światła, skoro można bardziej bezpośrednio zmierzyć absorpcję światła poprzez pomiar światła przepuszczanego i padającego. Zrozum więc, dlaczego LAI jest zmienną, którą jesteś zainteresowany.
Zastanów się również, czy LAI jest jedyną zmienną, którą chcesz zmierzyć. Widzieliśmy, że fotografia hemisferyczna może generować kilka wskaźników dotyczących struktury korony oprócz LAI, które mogą być przydatne. Czy pracujesz z wysokim czy niskim baldachimem? Jeśli pracujesz z bardzo wysokim baldachimem, być może nie jest możliwe umieszczenie nad nim czujnika NDVI , ponieważ po prostu nie masz infrastruktury, aby dotrzeć do szczytu baldachimu. W takim przypadku może być potrzebna fotografia półkulista lub pomiar przepuszczalności światła, taki jak LP-80.
Czy musisz mierzyć LAI dla poszczególnych gatunków? Jeśli tak, zbiór bezpośredni jest prawdopodobnie jedyną odpowiednią metodą. Czy chcesz przeprowadzać ciągłe czy dyskretne pobieranie próbek? Innymi słowy, czy chcesz, aby pomiary światła przechodzącego były stale rejestrowane, abyś mógł stale szacować zmiany LAI? A może wystarczy próbka punktowa? Załóżmy, że chcemy porównać LAI między różnymi poletkami. Być może podejście polegające na pobieraniu próbek punktowych jest bardziej odpowiednie.
Czy konieczne jest skalowanie pomiarów? Zastanów się nad protokołem pobierania próbek i dostępnymi źródłami danych, aby skalować od poziomu lokalnego do szerszej skali. Zastanów się, jak przestrzennie niejednorodny jest LAI w obrębie korony drzewa i jak bardzo jest on zbity. Będzie to miało wpływ na liczbę zebranych próbek i ich przestrzenne rozmieszczenie.
I wreszcie, czy potrzebne są bezwzględne wartości LAI, czy też można użyć wskaźnika zastępczego, takiego jak NDVI?
Pytania?
Nasi naukowcy mają wieloletnie doświadczenie w pomaganiu badaczom i hodowcom w pomiarach kontinuum gleba-roślina-atmosfera.
Wszystko, co musisz wiedzieć o pomiarze potencjału wody - czym jest, dlaczego jest potrzebny, jak go mierzyć, porównania metod. Ponadto można go zobaczyć w akcji, korzystając z krzywych uwalniania wilgoci z gleby.
Uproszczone zarządzanie nawadnianiem. Doskonałe zarządzanie wodą i składnikami odżywczymi bez straty czasu i pieniędzy z powodu problemów spowodowanych nadmiernym nawadnianiem.
: The researcher’s complete guide")