Leaf area index (LAI):研究者のための完全ガイド
葉面積指数は単一の数値であり、ある特定の時点における樹冠の統計的なスナップショットです。しかし、この一つの数値から重要な洞察が得られる可能性があります。
 and PRI (photochemical reflectance index)—The researcher’s complete guide")
現代の技術により、地球の表面全体をサンプリングする衛星から、個々の植物や葉を測定する携帯型の小型センサーまで、空間と時間の両方でさまざまなスケールにわたって NDVI や PRI などのスペクトル植生指数をサンプリングすることが可能になりました。

NDVI(正規化植生指数)とPRI(光化学反射率指数)はどちらも、電磁スペクトルにおける比較的狭い波長(10~50ナノメートル)の反射光を測定することで得られるスペクトル植生指数です。これは、植物群落の様々な特性を測定するのに役立ちます。NDVIは正規化植生指数、PRIは光化学反射率指数の略です。

スペクトル植生指標には多くの種類がありますが、この記事とウェビナーでは、最も一般的に使用される2つの指標であるNDVI とPRIの理論、手法、応用に焦点を当てます。
NDVIは、葉面積指数、光遮断率、さらにはバイオマスや成長といった植物群落の構造特性の測定に特に有用です。一方、PRIは光利用効率といった植物群落の機能特性の測定により有用です。最近の文献では、PRIが葉の色素の測定にも有用であることが示されています。
NDVIとPRIの起源を理解するには、樹冠と放射の相互作用について学ぶことが重要です。電磁放射が植物の樹冠と相互作用する際、主に3つの結果が存在します。

図2では、エネルギーは矢印の形をした光子として表されています。太陽から来る光子は、3つの方法で樹冠と相互作用します。透過、つまり樹冠を通過して地面に当たる光子。吸収、つまり樹冠内の緑色の光合成物質が光子の一部を吸収し、そのエネルギーを使って光合成を促進する。そして反射、つまり光は樹冠の表面に当たり、宇宙空間へと反射されます。

図3は電磁放射を示した図です。この図で最も顕著な特徴の一つは可視スペクトルです。可視スペクトルは400~700ナノメートルの範囲で、人間の目で見える青、緑、黄、オレンジ、赤をカバーしています。しかし、非常に短い波長の紫外線(UV)や、より長い波長の赤外線も存在します。電磁放射スペクトルには、異なるエネルギー(波長)の光子が存在することを理解することが重要です。植物の樹冠と放射線の相互作用は波長に依存します。例えば、赤色光子は青色光子や近赤外線光子とは異なる方法で樹冠と相互作用します。植生指数は、放射線の反射成分を測定します。
図 4 は、緑色の光合成物質が存在するほとんどの樹冠で見られると予想される典型的な樹冠反射スペクトルを示しています。y 軸は電磁放射の反射率 (%) を表します。x 軸はナノメートル単位の波長です。450 ~ 700 ナノメートルが可視光線範囲で、約 700 ナノメートルで赤外線、特にスペクトルの近赤外線部分に入ります。このスペクトルは植物の樹冠から得られたものです。可視スペクトルで反射される放射の大部分が緑色にピークを呈していることに注意してください。そのため、植物の樹冠は一般的に緑色になります。この場合、青と赤はクロロフィルによって強く吸収されます。また、近赤外線範囲の反射率が高いことにも注目してください。このエネルギーは光合成に使用できないため、この領域で高い反射率が見られるのは極めて一般的です。植物は、この放射を反射するように設計されています。

植物群落を構成する様々な構成要素は、可視光、近赤外線、そしてそれ以上の波長域にわたるスペクトルに影響を与える可能性があります。図5は、様々なレベルにおける葉のクロロフィル含有量を示しています。

クロロフィル含有量が変化すると、このスペクトルは、特に緑と赤の間の可視領域と近赤外線の少しの範囲で劇的に変化します。

同様に、図 6 は、カロテノイド含有量によっても反射率が変化することを示していますが、その影響はスペクトル全体にわたってそれほど広範囲には及んでいません。

葉の細胞構造もまた、樹冠の特性の一つであり、反射スペクトルに影響を与える可能性があります。図7は、その影響が可視光線と近赤外線のほとんどの波長域にわたって均一であることを示しています。

葉の水分含量も葉のスペクトルに大きな影響を与えます。図8は近赤外線スペクトルをより詳しく示しています。これは、水分の影響が最も大きい領域だからです。可視光域では影響は見られず、近赤外線域でもわずかな影響が見られる程度です。しかし、800ナノメートル付近では大きな影響が見られます。つまり、水分含量が影響を与えるのは、より長波長側なのです。
これまでの例は葉レベルでの影響を示しましたが、樹冠から観測される反射スペクトルは多くの葉が混在しています。これらをすべてまとめると、図9に示すように、葉面積指数もスペクトルの可視光域と近赤外域全体にわたって劇的な影響を与えていることがわかります。

約 600 ナノメートルから 700 ナノメートルの赤色領域と近赤外線領域では、葉の材料が追加されているため、反射率がどんどん高くなっていることに注意してください。
前のグラフは連続反射スペクトルを示していますが、NDVIはスペクトルの特定の2つの領域にのみ焦点を当てています。約800ナノメートルの近赤外線(NIR)と、通常約650~700ナノメートルの赤色領域の反射率を組み合わせたものです(図10)。典型的なNDVI値は-1~1の範囲です。NDVI値が高いほど、樹冠の緑が濃い、または葉面積が大きいことを示します。

前述のように、反射率は湧昇放射または樹冠から反射される放射と、樹冠に入射または当たるエネルギー量の比率です。NDVI が赤色と近赤外線のバンドで定式化される理由は、赤色がクロロフィル含有量に強く影響され、近赤外線が葉の細胞構造と葉内の空気空間に関係するためです。葉が拡大して成熟するにつれて、細胞構造が変化し、それが近赤外線の散乱に強い影響を与える可能性があります。また、図 10 のスペクトルから、赤色が強く吸収されていることもわかります。これは、あまり反射されないことからわかります。ただし、近赤外線では、反射率が大きくなっています。したがって、赤色と近赤外線の比率 (式 1) を取ることで、植生の樹冠で何が起こっているかを理解できます。

NDVIは、様々な樹冠の葉面積指数(LAI)を推定するためによく用いられます。これは、NDVIを経時的に追跡する時系列分析において特に有用です。葉面積指数の季節変動が大きい一年生落葉樹林冠では、NDVIはこうした変化を追跡するのに有用です。また、NDVIは葉面積指数の空間的変動を推測または推定するためにも使用できます。

図11は、研究者が画像を用いてLAIの空間変動を理解する方法を示しています。この画像には近赤外線と赤色波長のバンドが含まれているため、NDVIを計算できます。そして、得られたNDVI値を用いて作物のLAIをマッピングすることで、各管理単位間および管理単位内におけるLAIの空間的不均一性を把握することができます。
NDVI のこの応用には制限があることに注意してください。

図12は、LAIが3~4を超えると、NDVIと葉面積指数の関係が飽和する傾向があることを示しています。これは、赤色帯域では、LAIが低くてもクロロフィルの吸収が非常に低い傾向があるためです。そのため、葉面積が増えても、LAIが3または4を超えると吸収の変化はわずかになります。同様に、近赤外線帯域では、葉の材料が増えると、非常に高いLAIで近赤外線反射率の増分変化が減少する傾向があります。したがって、これはNDVIを使用してLAIを推定する際の潜在的な弱点の1つです。この方法は、LAIが0~4の範囲の樹冠にのみ適しています。
NDVIのもう一つの関連用途は、光吸収量の推定です。LAIは植物の樹冠が吸収する光量と関連しているためです。葉の数が多いほど、吸収される光量も多くなります。光吸収量は、樹冠が吸収する光の量をある程度把握できるため、重要な変数です。例えば、樹冠が環境に順応し、その光エネルギーをすべて利用できると仮定すると、光吸収率を知ることで光合成能力をある程度把握できます。
光合成有効放射(FPAR)の分光吸収率または分光吸収率を予測するためにNDVIを使用する利点の一つは、NDVIとFPARの関係が通常飽和しないことです。光吸収は、LAIが約3~4のときに比較的完全に完了する傾向があります。葉面積が増加しても、分光吸収率は直線的に増加しません。図13は、NDVIとFPARの直線関係を示しています。

分数インターセプトの値が高くても、曲線は直線的です。
NDVIの比較的一般的な用途として、落葉樹や一年草、あるいは葉面積の年次変動が大きいその他のタイプのシステムにおけるフェノロジー(季節変化)の推定があります。図14の下のグラフは、NDVIデータの時系列データに曲線を当てはめると、かなり規則的なパターンを示すことを示しています。

冬季は葉面積が広いだけでなく、NDVIも低くなります。春になると、LAIの増加に伴いNDVIも増加します。生育期の中頃にピークに達し、秋には葉の老化と落葉によりNDVI値は低下します。これらの曲線を用いることで、様々なイベントのタイミングに関する様々な指標を抽出できます。例えば、緑化や季節の始まりのタイミングに関心がある場合を考えてみましょう。図14の上のグラフは、NDVI曲線から抽出されたSOSp指標を示しています。このSOSp指標は、葉が成長し始めた時期を示しています。葉が成長し始めた時期がわかるため、特定の日付を季節の始まりとしてマークすることができます。このような観測結果を長年にわたりまとめると、特定の樹冠において季節の始まりが発生した日付が多数得られます。これを気候変数やその他の変数と関連付けることで、因果関係があるかどうかを調べることができます。例えば、一部の研究者は、気候が温暖化すると、開花期が年内でより早く訪れることを発見しています。また、これらの曲線から抽出できる指標は他にも数多くあります。NDVIデータをフェノロジーの指標として直接使用することも、フェノロジーイベントの発生時期を予測するモデルに直接入力することもできます。

図15は、NDVIを用いてフェノロジーを理解する別の例を示しています。混合種が生息する林分では、樹冠内の様々な場所、あるいは個々の樹木を観察し、それぞれの成長時期を追跡することができます。図15は、春季における混合種林の発達を示しています。すべての樹種は、当初は非常に異なるNDVI値を持っています。これは、その時点における葉面積指数に直接関係しています。春が進むにつれて、落葉樹のNDVI値は通常増加し、その後、約150日目以降、ほぼ均一なNDVI値に収束する傾向があります。つまり、約150日目には、樹冠は構造的に成熟し始めていることになります。この時点で光合成順応がどのように進行しているかは必ずしも明らかではありませんが、これらのデータは、NDVIデータを用いて空間的な異質性、あるいは同一林分内であっても、樹木ごとの緑化の違いを評価する方法の良い例となっています。
NDVI データは、緑葉面積の存在が季節的な光合成と非常に密接に結びついている一年生草地などの、落葉樹林冠または季節性が強い林冠における生産性を直接推定するために使用できます。

例えば、図 16 は Ryu 氏の論文のデータで、彼は数年にわたって一年生草原の NDVI を追跡しました。NDVI 値は上のグラフにプロットされた緑の点で、同じグラフの白丸は光合成の測定値です。NDVI は、この一年生草原における光合成のタイミングと振幅の追跡に優れていることに注目してください。下のグラフでは、彼は 4 年間のデータセット全体を取得し、NDVI と樹冠光合成の相関関係を示しています。したがって、彼はこの生態系内の樹冠光合成を推定するために NDVI を使用する経験的モデルを作成しました。NDVI はこのシステムの光合成のタイミングを追跡するため、年間の任意の日の瞬間光合成を推定できるだけでなく、「炭素更新期間をどのように推定できますか?」「シーズンはいつ始まりましたか?」「ピークはいつですか?」「いつ終わりましたか?」などの質問に答えることもできます。図 16 は、さまざまな有用な変数をマイニングできる豊富なデータセットです。
NDVI 、考慮すべきいくつかの制限がある:

図 17 は、1 年間を通して NDVI 値がかなり安定しているのに対し、樹冠の CO2 吸収または光合成は時間の経過とともに動的であることを示しています。しかし、NDVI の時間的ダイナミクスと樹冠の CO2 吸収の間には関連性がありません。したがって、これは NDVI の不適切な使用法です。私たちは、樹冠について、そして機能的変化の原因についてもっと知りたいのです。そうは言っても、NDVI は常緑樹システムにおいて空間的異質性を調べるのに役立ちます。広い領域または長期にわたる LAI または光遮断バイオマスの変動を調べる場合です。たとえば、山火事後の回復と成長を評価する場合、たとえそれが常緑樹システムであっても、十分に広い期間を調べると、NDVI はシステム内で反応します。ただし、年間のタイムスケールでは、おそらくそれほど動的な変化はないでしょう。
PRIはNDVIと計算式が似ていますが、入力として使用される波長またはバンドのみが異なります。PRIは531ナノメートルと570ナノメートルの反射率で計算され、PRI値の範囲は-1から1です。

光化学反射率指数は、植物群落内で発生する光利用効率、あるいはその変化を調べるために最も一般的に用いられます。これは、特にキサントフィル色素の変化に対する531ナノメートル波長の応答を調べることによって行われます。

図18は、比較的短い間隔で連続して取得した2つの異なる反射スペクトルを示しています。これら2つのスペクトルの差を計算すると、差が最も大きいピーク領域がいくつか見つかります。そのうちの1つ、約531ナノメートルのピークは、キサントフィルサイクルの活性に起因すると考えられます。
キサントフィルサイクルは、キサントフィル色素が前方と後方の両方向で内部変換プロセスを経る、迅速かつ可逆的な作用/反応です。

図19は、その内部変換プロセスを示しています。図の右側では、ビオラキサンチンから始まり、アンテラキサンチンに変換され、最終生成物はゼアキサンチンです。ビオラキサンチンはストレスのない状態であり、植物がストレスに遭遇すると、色素の内部変換が起こり、ゼアキサンチン状態の色素が増加します。ストレスが解消されると、このプロセスは可逆的です。例えば、光レベルを下げると、これらの色素の内部変換は逆転し、植物組織内の色素の大部分はビオラキサンチンになります。これは、植物がストレスの多い環境に安全に対処するための普遍的な方法です。植物は屋外にいるとき、絶えず放射線を吸収しています。立ち上がって立ち去ることはできません。このプロセスは、植物が余分な光をキサントフィルサイクルに吸収する方法であり、そのエネルギーは光合成機構に悪影響を与えるのではなく、色素の内部変換を促進するために使用されています。光が吸収され、キサントフィルサイクルに利用されると、光利用効率が低下します。これは、光が炭素吸収を促進せず、代わりにキサントフィルサイクルを介した熱放散プロセスに利用されるためです。したがって、キサントフィルサイクルは、植物が光利用効率を制御する方法の一つです。
キサントフィルサイクルは、時間経過に伴う他の興味深いダイナミクスも示します。例えば、デミッグ、アダムス、アダムスによる論文の図20は、いくつかの異なる処理と、キサントフィルプールの総量(ビオラキサンチン、アンテラキサンチン、ゼアキサンチンの含有量の合計)の予測を示しています。また、相対的なプールサイズ、つまりビオラキサンチン、アンテラキサンチン、ゼアキサンチンの比率も示しています。

上の円グラフは、日陰の葉と日向の葉を比較したものです。日向の葉では、プールの総量が日向の葉よりもはるかに大きくなっています。各キサントフィルの割合に注目してください。日陰の葉では、キサントフィルの多くがビオラキサンチン状態にあり、これはおそらくサンプル採取時点では光保護の必要性が低いためと考えられます。一方、日向の葉では、キサントフィルの大部分がゼアキサンチン状態にあり、光保護が必要であることを示しています。
夏季と冬季のキサントフィル含有量を比較した場合も同様のことが見られます。例えば、冬越しする常緑樹では、夏季は光保護の必要性が大幅に減少します。これは、植物が吸収するエネルギーの大部分(ストレスがない場合)を光合成に利用できるためです。しかし、冬季は気温が低く、光合成に適さない条件となります。植物は依然として光を吸収していますが、自己破壊を防ぐために何らかの形で光を利用しなければなりません。そのため、植物はキサントフィルのプールサイズを増やすことに資源を投入します。つまり、冬季には光保護の必要性が高まるため、ゼアキサンチンの割合が増加します。
下の例では、窒素処理に違いがありますが、基本的には他の2つの例と同じパターンです。植物に窒素が豊富に含まれている場合、ストレスが少なく、光保護の必要性も低くなります。窒素が少ない場合、吸収した光子を利用できず、キサントフィル色素による光保護の必要性が高まります。
研究者がPRIを利用する方法の一つは、キサントフィルの時間的ダイナミクス(すなわち、キサントフィルの活性による光利用効率)を捉えることです。PRIの利用は、ジョン・ガモン博士が反射スペクトルを用いてキサントフィルサイクルの活性を検出できることを発見したことに始まります。彼は、531ナノメートルの反射率がキサントフィルの変化に最も敏感であることを発見しました。

図21の右上は531ナノメートルにおける反射率を示しており、X軸の酸化状態は、ビオラキサンチン、アンテラキサンチン、ゼアキサンチンの比率で表されたキサントフィルサイクルの状態を示しています。この短い時間間隔で、キサントフィルサイクルのダイナミクスと、531ナノメートルにおける反射率がそれらの変化に敏感であることがわかります。
PRIは570ナノメートルのリファレンスバンドも使用している。キサントフィルが変化するときに570ナノメートルで発生する変化はありません。

図22は、PRIの変化がキサントフィルの酸化状態の変化に対して、531ナノメートルにおける反射率とほぼ同程度に敏感であることを示しています。また、キサントフィルの活性は反射スペクトル内の比較的狭い帯域内でしか検出できないことにも注意してください。この場合、広い帯域(つまり、緑色スペクトル全体にわたる測定)では、キサントフィルサイクルの活性を見逃す可能性があります。したがって、キサントフィル関連の変化を検出する、あるいは検出できるほどの感度を持つには、狭い波長帯域での測定が必要です。
一部の研究者は、PRIを用いて植物の光合成機能の日周ダイナミクスや短期ダイナミクスを解析しています。図23と24は、PRIが光利用効率の変化にいかに動的に反応するかを示す簡単な実験例を示しています。

図23は、研究者が植物に段階的に光を照射した実験です(光レベルはピークに達するまで増加し、その後低下します)。上のグラフは、光レベルが段階的に上昇し、その後低下する様子を示しています。また、CO2吸収量(光合成を示す)も光レベルの変化に追従しています。つまり、利用可能なエネルギーが増えると光合成が増加し、光レベルが減少すると光合成は減少します。
図23の下のグラフは、全く同じ実験で異なる測定値を用いたものです。2つの測定値はPRIと、光化学系IIの効率(光利用効率に関連)を測定する蛍光測定値であるΝF/Fmです。図24は、PRIと蛍光測定値が光レベルの変化のタイミングを良好に追跡していることを示しています。ここでも、光レベルが上昇するとPRIとΝF/Fmが減少し、どちらも光利用効率の低下を示しています。そして、光レベルが下降すると、その逆の現象が見られます。

図24は、蛍光とPRIの経時的な変化を示すもう一つの簡単な実験です。植物は0時間目に暗闇の中で光から光へと変化します。1時間後、光が点灯すると、蛍光とPRIはほぼ即座に反応を示します。光利用効率と相関して、非常に強い下方反応を示します。その後、しばらく光が点灯した後、実験者は光を消します。すると、葉の生理学的変化に伴い、光量の変化に対して蛍光とPRIの両方が比較的迅速かつ即座に反応します。
これらの実験は、PRIが一日を通して起こり得る急激な変化や、植物の樹冠が日向から日陰へと変化する際の変化を測定するのに役立つ可能性を示しています。これらの変化やその他の環境変数が変化すると、PRIが検出できる可能性のある光利用効率の変化が期待されます。
PRIを長期測定に利用するというアイデアは以前から存在していましたが、長期間にわたるPRIの測定はやや困難であったため、この分野を追求する研究者はそれほど多くありませんでした。通常、PRIのような狭い波長の光を得るには高度な機器が必要です。しかし、近年関心が高まっており、図25に示す結果は比較的新しいものであり、現在も研究が続けられています。
研究者らがこれまでに発見したのは、長期的には、PRI はキサントフィルサイクルに敏感であるだけでなく、総カロテノイドとクロロフィル含有量にも敏感であるということです。キサントフィル(ビオラキサンチン、アンテラキサンチン、ゼアキサンチン)はカロテノイドの一種であることに注意することが重要です。そのため、研究者が総カロテノイド含有量を測定すると、その大部分は実際にはキサントフィルであることがよくあります。PRI がキサントフィルだけに反応しているのか、総カロテノイドに反応しているのかはまだ完全には明らかではありません。さらなる研究がそれを明らかにするのに役立ちます。しかし、研究者らは、PRI とカロテノイドとクロロフィルの比率の間に非常に密接な相関関係があることを発見しました。カロテノイドは補助色素であり、集光役割を果たすことができるため、カロテノイドとクロロフィルの比率は光利用効率と関連付けることができます。しかし、キサントフィルをはじめとする多くの色素は、光保護の役割も担っています。そのため、カロテノイドとクロロフィルの含有量を比較することで、その比率の経時的な変化を観察し、植物がどの程度ストレスを受けているかを把握することができます。つまり、PRIを用いて光利用効率や光合成効率をより広い期間や広い空間領域で評価する方法となる可能性があります。

図25の左側のグラフは、ヨーロッパアカマツの樹冠を1年強観察し、光合成生理、葉の色素、およびPRI間の長期的な相関関係を評価した研究を示しています。この研究では、重要な光合成生理学的変数のほぼすべてがPRIと関連していることがわかりました。これは、これらの変数間に多くの共相関関係があることが一因ですが、先行研究と一致して、キサントフィル対クロロフィル比、カロテノイド対クロロフィル比、そしてキサントフィルの総プールサイズと総カロテノイドプールサイズとの間に最も高い相関関係がいくつか見られました。
しかし、多くの共相関関係があるにもかかわらず、図 26 を見ると、研究者は依然として PRI を長期間にわたって使用して光利用効率の傾向を推定できることを示しています。

このように、PRIは日周的な時間スケール全体にわたって光利用効率を測定するのに優れており、より長い時間スケールでも使用できるようです。しかし、より長い時間スケールで使用する場合、PRIの変化を引き起こす駆動要因を真に理解するためには、データの解釈に一定の注意を払う必要があります。
NDVIと同様に、PRIにもいくつかの限界があります。まず、PRIと光利用効率の関係は、時間スケールに関わらず、樹冠の種類によって異なることが分かっています。つまり、樹種や構造によっても異なるのです。

図27は、多くの異なる種において、PRIと蛍光(光利用効率の代理指標)の関係が種によって異なることを示しています。そのため、PRIを測定して光利用効率の値を抽出することはできません。絶対的な光利用効率の値を得るには、まず相関関係を構築するための背景作業を行う必要があります。これはNDVIにも当てはまります。NDVI値が分かっている場合、そのNDVI値を絶対的なLAI値またはFPAR値に変換する前に、相関関係を構築するための作業を行う必要があります。
生育期全体を通して、PRIは多くの変数の変化に反応しますが、そのほとんどは光合成能力に関連しています。しかし、ある期間のPRI測定値を別の期間の測定値と直接比較する場合、また、これらの測定値の一部を空間的にスケーリングする場合は注意が必要です。
また、PRIと光利用効率は、特に極端な場合には切り離される傾向があることにも留意してください。例えば、常緑樹の生態系では、春から夏への移行期にしばしば切り離しが見られます。研究者が切り離しを観察したもう一つの事例は、極度の干ばつ時です。樹木が枯死寸前になると、PRIは機能不全に陥る傾向があります。このような状況では、多くの場合、PRIは光利用効率を過大評価してしまうことになります。したがって、繰り返しになりますが、注意が必要です。
最後に、光利用効率はキサントフィルサイクル色素やその他の葉の色素含有量だけに関連するのではありません。場合によっては光呼吸も考慮する必要があり、PRIは光呼吸とは無関係です。したがって、PRIに基づく光利用効率の推定において、光呼吸は誤差の原因となる可能性があります。
光合成を遠隔的または非破壊的に推定できることに、幅広い関心が寄せられています。一部の研究者は、モンティスの光利用効率モデル(式3)に立ち返っています。

ここで、GPP(総一次生産性)または光合成は、植物の樹冠に照射されるPAR(光合成有効放射)とFPAR(光吸収率)の積、つまり、そのPARのうち実際に植物の樹冠に吸収される量とε(光利用効率)の積です。言い換えれば、吸収された光子は、炭水化物の形で炭素を固定するためにどれほど効率的に利用されているかということです。
しかし、一部の研究者は FPAR と ε に対してはプロキシを使用することを提案しているものの、PAR に対してはプロキシを使用しないことを提案しています。

研究者たちはFPARの代わりにNDVIを、εの代わりにPRIを代用する試みを行っており、良好な結果が得られています。図28は、時間と空間の両方にわたってスケーリングした研究です。

各データポイントは、異なる場所、または同じ場所で異なる期間にサンプリングされたものです。この場合、NDVIにsPRIを乗じただけです(sPRIは、PRIの値が0と1の間になるようにスケーリングする方法です)。sPRIは基本的にPRIと等価であり、図28からわかるように、その関係はかなり緊密です。

図29は、PARとFPARをAPARにまとめたものです。APARは吸収された光合成有効放射です。つまり、PARとFPARを組み合わせ、さらにスケール化されたPRIを掛け合わせたようなものです。これらの研究者は、実際の光合成単位で光合成を推定しようとしています。ここでも、かなり直線的な関係が見られます。スケールに関しては多少のバイアスはありますが、光合成を遠隔的に検出するという点では非常に優れた成果です。
NDVI と sPRI 光利用効率型モデルを組み合わせた 1 つの用途は、空間スケーリングです。

図30は衛星画像から得られた結果です。この衛星画像から、研究者はNDVIとPRIを計算しました。彼らは光利用効率モデルにNDVIとPRIを組み合わせ、画像内の各ピクセルまたは各位置について、CO2吸収量または光合成量の推定値を抽出しました。これは衛星画像から得られたものですが、地形全体に多数のセンサーを配置することで、非常に類似した結果を導き出すことができる、あるいは少なくともこのNDVI/PRIを組み合わせたアプローチを用いて光合成における空間的不均一性を把握することができると考えられます。
測定できる光の波長が多様であるため、スペクトル データを測定する方法は多様です

図31は、様々な機器による様々な測定値を示しています。例えば、分光計から得られる「ハイパースペクトルデータ」と呼ばれるデータがあり、スペクトル全体にわたってほぼ連続的な情報が得られます。これは、機器から抽出できる最も詳細なスペクトルデータと言えるでしょう。通常、この種の機器は、新たな植生指標や新たな動態の探索といったリモートセンシング研究用途で使用されます。例えば、PRIは複数のスペクトルを一度に探索することで発見されました。
もう一つのアプローチは、限られた数のスペクトルバンドを使用することです。例えば、QuickBird衛星やその他の高解像度衛星からは、「マルチスペクトルデータ」と呼ばれるデータが提供されます。QuickBirdは、青、緑、赤、近赤外線の4つのバンドを提供します。これらのバンドだけで、この領域における植物群落のスペクトルダイナミクスを表現できます。つまり、マルチスペクトルアプローチは、基本的にマルチバンド放射計と同義です。ハイパースペクトルとマルチスペクトルの違いは曖昧ですが、基本的に「ハイパー」は多くのことを意味し、「マルチ」は少数のことを指します。極端な例としてハイパースペクトルがありますが、その反対にはシングルバンド放射計があります。PAR機器は、基本的に非常に広い波長帯域を測定できるシングルバンド放射計です。400~700ナノメートルの放射を測定し、その領域全体に対して単一の値を出力します。日射計も同じことを行いますが、その測定範囲はさらに広く、近赤外線および短波赤外線領域までずっと広範囲にわたります。
したがって、必要な機器は研究目的によって決まります。もしこの記事で紹介した特定の用途の測定だけを目的とするのであれば、フル機能の分光計は過剰でしょう。そのような場合には、PRI、NDVI、あるいはその両方を同時に測定できるマルチバンド放射計を使用できます。必要なのは4つのバンドだけです。
METER のNDVI/PRI センサーは、NDVI と PRI の両方を測定するマルチバンド放射計です。

1つは630ナノメートルと800ナノメートルを中心とする波長帯域を持ち、NDVI測定用に設計されています。もう1つは532ナノメートルと570ナノメートルを中心とする波長帯域を持ち、PRI測定用に設計されています。分光反射率センサーは入射光を測定するように設計されています(図33)。

テフロン製の拡散板は、空を半球状に観測し、入射光の測定を可能にします。右下には、視野を20度に制限する物理的な視野絞りを備えた方向センサーが配置されています。また、このセンサーは方向を指示できるため、研究者は植物の樹冠内でセンサーがどの方向を見ているのかを実際に指示することができます。NISTトレーサブルな校正が施されているため、測定値と実際の光の物理単位を提供します。小型で防水性と耐候性を備えているため、屋外に設置して長期間にわたってデータを収集する必要がありません。これは、多くの分光計で従来問題となっていた点です。通常、この機器を風雨から保護するために、何らかの容器を用意する必要があります。
PRI と NDVI を取得するためにスペクトル反射率を測定する場合、通常は、少なくとも放射輝度測定 (または湧昇測定) については、樹冠を上から見たビューが重要になります。

これには、樹冠上方からの測定が必要です。この作業の難易度は樹冠の高さに依存します。例えば、図34に示すように、ミシガン州北部には、混合落葉樹林のかなり上まで伸びたフラックスタワーがあります。これはSRSセンサーに最適なプラットフォームです。しかし、例えば小麦畑のような樹冠であれば、タワーは不要です。センサーをフェンスの支柱や頑丈な三脚に設置できます。いずれにしても、センサーは樹冠より上方に設置する必要があります。
分光反射率を長期にわたって測定する場合、デュアルビュー機器を使用すると便利です。1 つの機器は上を見上げて入射放射を定量化し、もう 1 つの機器は下を見下ろしてその放射の反射量を定量化します。つまり、空の状態は関係なく、日光が当たる状態で測定でき、雲が流れてきても両方の放射源が同時に定量化されるため測定に影響しません。これは、衛星画像ではなく地上ベースの分光反射率測定を行うよりも有利な点です。衛星画像の問題は、特に可視領域で雲が覆っているときはいつでも、対象の樹冠の生態系の視界が遮られることです。しかし、あらゆる条件下で樹冠を測定できる SRS センサーまたは分光計を使用すると、あらゆる条件下で NDVI と PRI をより詳細に調査できます。また、キャノピーの上にセンサーを設置する場合は、キャノピーを継続的に監視できる場所に設置するのが簡単なので、小型で低電力のセンサーを使用するのが有利です。
DVIとPRIに関して留意すべき点の一つは、太陽センサーの表面形状の変化がデータに劇的な影響を与える可能性があることです。これは時系列データを検討する場合に特に重要です。例えば、図35は5日間のNDVIをプロットしたものです

それぞれの線は異なる日を表しています。これらのデータ線の中に、典型的なU字型のパターンがあることに注目してください。これは角度の影響であり、変化していたのはLAI(光子捕捉率)ではなく、光子が樹冠とどのように相互作用し、機器によってどのように記録されるかに関係しています。
図 36 は後方散乱と前方散乱の極端な例を示しています。

この例では、観測者と樹冠の位置は変化していませんが、空における太陽の位置は変化しています。これは、光子が樹冠にどのように当たるか、そして観測者や機器の位置で光子がどのように記録されるかに非常に大きな影響を与える可能性があることがわかります。
また、あらゆる気象条件下で長期にわたる時系列データを収集しているため、観測結果に誤差が生じる可能性がある点にご留意ください。前部光学系が濡れたり、鳥がセンサーに止まったり、センサーが汚れたりする可能性があります。こうした状況はすべて、データにノイズやスパイクを引き起こす可能性があります。例えば、図35の178日目には、大きな上昇スパイクが見られます。

そのため、時系列の NDVI および PRI データでは通常、ある程度のデータ フィルタリングが必要です。太陽センサーの表面形状の影響に対する解決策の 1 つは、1 日に 1 つの観測値のみを使用することです。NDVI は LAI など、1 日を通してあまり変化しないが、日ごと、週ごと、月ごとに変化する構造的に関連した変数を調べるため、この方法は通常 NDVI ではうまく機能します。そのため、この点に焦点を当てる場合は、日ごとの解像度だけで十分です。図 37 は、研究者が毎日正午頃に 1 つの測定値を抽出した例です。観測データが取得されるとき、太陽の天頂と方位は日ごとに一定であるため、正午、特に太陽の正午が適しています。
図 38 は、いくつかの異なる処理区からの時系列であり、草地の春から夏への移行期の緑のパターンまたはパターンの違いを示すために毎日のデータがプロットされています。

一部の研究者は日周データを求めており、特にPRIはキサントフィルサイクルのダイナミクスに反応するため、非常に動的である可能性があります。植物や植物群落が環境の急速な変化に反応する際に、これらの緊密な連携と迅速な反応がどのようなものなのかを理解することには、大きな関心が寄せられています。日周データを使用する場合は、2008年にRemote Sensing of Environment誌に掲載されたThomas Hillkerの論文に記載されている双方向反射率モデリング(データに存在する角度効果をモデリングまたは考慮する手法)を検討することをお勧めします。
当社の科学者には、研究者や生産者が土壌-植物-大気の連続体を測定するのを何十年も支援してきた経験があります。
このビデオを視聴するか、以下の記事を読んで、NDVI が現在の研究でどのように使用されているか、また NDVI の制限の一部を克服する方法を学びましょう。
NDVIを最も広く捉える方法は、地球を周回する衛星から得られるデータです。図1では、地球全体で濃い緑色で表された、植生が豊かでNDVI値が高い地域が確認できます

逆に、植生の少ない地域ではNDVI値が低く、茶色く見えます。NDVIは、地球表面全体に存在する植生の被覆量に敏感です。
NDVIは区画レベルでどのように役立つのでしょうか?下の図2は、遷移勾配を示しています。ここでは、時点0は裸地、または少数の雑草や一年草が生えている状態です。この土地を十分な時間放置すると、植生は変化します。低木が草地を覆い、最終的には森林が見られるようになるかもしれません。広い範囲では、草原から森林へと移行することもあります。農業システムでは、裸地から植物の出現、成熟、そして老化へと、植生は毎年入れ替わります。このサイクルは毎年繰り返されます。これらの成長サイクルの中で、NDVIは、時間の経過とともに起こる樹冠の成長と、景観全体で起こる空間的なダイナミクスを定量化するのに役立ちます。

では、NDVIはどこから来るのでしょうか?図3では、x軸は電磁スペクトル内の光の波長を示しています。450~950nmは可視光線領域と近赤外線領域の両方をカバーします。y軸は反射率(%)です。これは緑地からの典型的な反射スペクトルです。

緑色のハイパースペクトル線は、分光放射計から得られると予想されるものです。反射率は通常、青色領域では低く、緑色領域では高く、赤色領域では低くなります。可視光から近赤外線領域に移ると、反射率は劇的に変化します。NDVIと表示された2本の縦線は、典型的なNDVIセンサーがスペクトル内のどの領域を測定するかを示しています。1つの帯域は赤色領域にあり、もう1つは近赤外線領域にあります。
NDVIは、可視光線領域と近赤外線領域のスペクトルの大きな差を利用します。健全で成長している植物は近赤外線を強く反射します。上図の右側にある2つの画像は同じ領域を撮影したものです。上の画像はトゥルーカラー、つまり青、緑、赤の3つのバンドで表示されています。下の画像は擬似カラーの赤外線画像です。表示されている3つのバンドは青、緑、そして赤の代わりに近赤外線を使用しています。鮮やかな赤色は、緑または健全な植物に特徴的な近赤外線反射率が高いことを示しています。
NDVIが赤色と近赤外線を用いて算出されるのは、赤色がクロロフィルの吸収に関係し、近赤外線が樹冠構造と葉の内部細胞構造に敏感だからです。樹冠に葉が増えるにつれて、クロロフィルと構造の複雑さが増し、赤色の反射率が減少し、近赤外線の反射率が増加すると予想されます。

正規化植生指数(NDVI)は、植物による近赤外線(NIR)の反射量を考慮します。これは、近赤外線と赤色光の反射率(Rho)の差を、両者の合計で割ることで算出されます。NDVI値は通常、-1(地表水)から1(完全で活気のある樹冠)の範囲です。低い値(0.1~0.4)は樹冠がまばらであることを示し、高い値(0.7~0.9)は完全で活気のある樹冠であることを示します。
反射率を計算する方法は、湧昇放射(樹冠に当たり、その後センサーに向かって反射される放射)と、樹冠に降り注ぐ(上空から)放射の総量の両方を定量化することです。この2つの比率から、各バンドにおける反射率が得られます。
NDVIは、葉面積指数(LAI)や樹冠の分光受光率(FPAR)などを推測するために用いられます。科学者の中には、NDVIをバイオマスや作物の収量と関連付ける人もいます。また、NDVIはフェノロジー(緑化の一般的な時間的パターン)を把握したり、植生がどこに生息しているか、特定の場所にどれだけの植生が生息しているかを把握するためにも用いられます。
図 4 では、特定の樹冠 LAI における反射率スペクトルが葉面積指数によってどのように変化し、可視範囲では減少し、近赤外線では増加するかがわかります。

LAIが非常に低い場合、反射スペクトルは赤色光と近赤外線(黒線)の間で比較的区別がつきませんが、LAIが高い場合、クロロフィルによる赤色光の吸収が強く、近赤外線の反射率が高くなります。実際、LAIが増加すると、800 nm付近の近赤外線の反射率は増加し続けます。
正規化植生指数(NDVI)の限界は、スペクトルの両端で発生する傾向があります。植生被覆率が非常に低い場合(シーンの大部分が土壌)、NDVIはその土壌の影響を受けます。これにより、測定値が混乱する可能性があります。一方、植生が非常に多い場合、NDVIは飽和する傾向があります。葉面積指数(LAI)が3(紫)と6(緑)のスペクトルの違いがほとんどないことに注目してください。実際、熱帯林ではLAIが既に非常に高いため、NDVIはLAIの小さな変化には敏感ではありません。しかし、いくつかの解決策があります。
図5は、裸地の横断面全体にわたって異なる植生指標のスペクトル測定を行った研究の結果を示しています。乾燥した埴壌土から湿潤な埴壌土に移ると、土壌の湿潤度によってNDVIが強く反応することがわかります。これは植生を測定するには望ましくありません。土壌や土壌水分の変化に敏感な指標は対象としていません。しかし、図5には、横断面全体の土壌の変化に対する感度がはるかに低い指標がいくつかプロットされています。

これらの指標の1つは、土壌調整植生指数(SAVI)です。SAVIの計算式はNDVIに似ており、NDVIと同じ2つの波長帯、つまり近赤外線と赤色光が組み込まれています。


唯一の違いはLパラメータです。Lは土壌調整係数で、0から1までの値を取ります。植生被覆率が100%の場合、土壌背景調整は不要なため、Lは0になります。しかし、植生被覆率が非常に低い場合、Lパラメータは1に近づきます。NDVIを使用せずに植生被覆率を正確に測定することは難しいため、事前にLを推測することで、土壌の影響を受けないようにNDVIを調整することができます。Lは中間値である0.5に設定するのが一般的です。図5を見ると、土壌調整植生指数(SAVI)は土壌背景に対する感度がはるかに低いことがわかります。
次の植生指数は、修正SAVI(MSAVI)です。SAVI方程式には推定しなければならないLパラメータが含まれており、これは正確な処理方法とは言えません。そこで、Qiという科学者がLの普遍的な最適値を開発しました。ここでは数式には立ち入りませんが、彼はSAVI方程式を簡素化し、Lパラメータを不要にし、近赤外線と赤色の反射率のみを入力するだけで済むようにしました。


これは、L を推定したり独立して測定する必要がなくなったため、非常に重要な進歩でした。Qi が SAVI と MSAVI を比較したところ、植生量に対する感度と土壌背景に対する反応の点で、2 つの指標に実質的に違いはありませんでした。

NDVIは、土壌の影響が強くない限り、LAIの中程度の範囲で有用ですが、LAIが4を超えると感度が低下します。図7では、感度の低下は主に赤色帯域の飽和によるものです。測定は小麦の群落とトウモロコシの群落で行われました。近赤外線反射率は小麦とトウモロコシの群落の全スペクトルにわたって感度が高いですが、赤色帯域は比較的早く飽和します。赤色帯域が飽和し始める地点がNDVIの飽和地点です。

注:NDVIはLAIが高いと飽和しますが、光合成有効放射の分光吸収率(FPAR)を測定することが目的であれば、NDVIでは飽和の問題は発生しません。図8では、FPAR、つまり光合成有効放射の分光吸収率は、NDVIが飽和するずっと前にほぼ完了しています。これは、樹冠が効率的に光を吸収するため、LAIが約4になると、ほとんどの光が樹冠によって遮断または吸収されるためです。したがって、LAIを徐々に増加させても、FPAR変数には大きな影響はありません。

NDVI飽和問題に対する解決策の一つとして、広域ダイナミックレンジ植生指数(WDRVI)が挙げられます。その定式化はNDVIと似ていますが、近赤外線と赤色光の反射率の寄与の差を軽減するために重み付け係数が用いられます。

WDRVIでは、aに近赤外反射率を乗じて値を小さくし、赤色反射率の値に近づけます。そうすることで、植生指数に対する赤色と近赤外域の寄与をバランスさせます。

図 9 は、a の値が小さいほど、LAI に対する広域動的植生指数の線形応答が増加することを示しています。
WDRVIの唯一の欠点は、aの選択が主観的であることです。最適なaの値を見つけるまで、自分で試行錯誤する必要があります。aが減少するにつれてLAIへの線形応答に近づくため、人々は非常に低い値を選択する傾向があります。
強化植生指数(EVI)は、高バイオマス生態系における感度を高めるために設計されましたが、大気の影響も低減することを目的としています。これは、衛星ベースのプラットフォーム向けに作成された植生指数です。衛星から地上までは、大気を透過する層が多く、大気中のエアロゾルが赤色光および近赤外線領域の反射率に影響を与え、誤った観測結果をもたらすことがあります。EVIは、土壌に対する指数の感度を低減する効果も期待しています。つまり、EVIは両極端の問題に対する一種の解決策と言えるでしょう。

EVIの式では、近赤外線と赤色光の反射率が2つの主要な入力となります。C1、C2、Lはすべて推定可能なパラメータですが、青色帯域は測定が必要です。ほとんどのNDVIセンサーは2バンドセンサーであるため、青色帯域の情報は得られません。さらに、衛星データの場合、青色帯域は比較的ノイズが多く、必ずしも最高品質のデータが得られるとは限らないため、EVIの価値は限られています。
これらの問題から、Jiangという科学者が解決策を思いつきました。Jiangは赤いバンドと青いバンドの間にかなりの自己相関があることに気づき、青いバンドを除いたEVIをEVI2(Enhanced Vegetation Index 2)と名付け、定式化しようと試みました。数式にご興味があれば、彼の論文をお読みになることをお勧めしますが、ここでは、実際に使用する場合に備えて式をご紹介します。
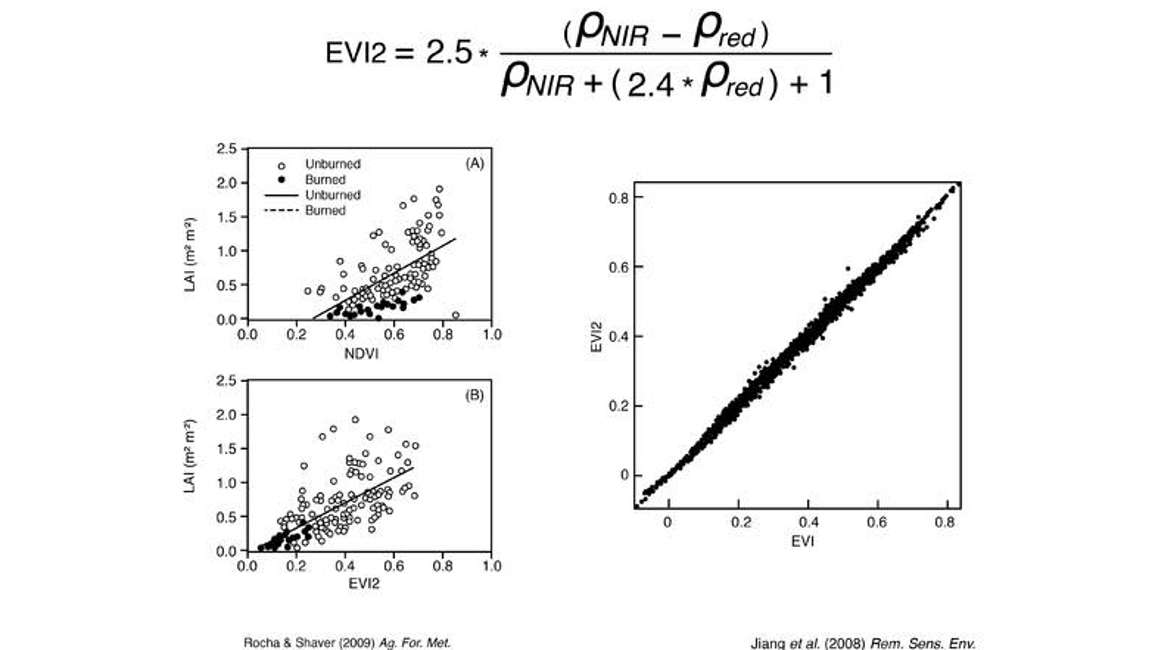
JiangがEVI2を計算し、従来のEVIと比較したところ(図10)、ほぼ1対1の関係が見られました。事実上、EVI2はEVIと同等でした。EVI2は青色バンドを回避しているため、近赤外線(NIR)と赤色バンドという2つの入力値のみを用いてNDVIを計算できるため、非常に興味深い可能性を秘めています。
NDVI測定は非常に価値があります。NDVIの性能が極端に低い場合もありますが、そのような場合でも複数の解決策があります。これらの解決策はすべて近赤外線と赤色のバンドを使用しているため、NDVIセンサーを使用して近赤外線と赤色の反射率の生の値を取得し、これらの指標のいずれかで再定式化することができます(ここで取り上げていない他の指標もいくつかあります)。したがって、LAIが極端に高い、または低いシステムを使用している場合は、近赤外線と赤色のバンドを何らかの植生指標にどのように使用できるかを検討し、特定のアプリケーションを調査できるようにしてください。
当社の科学者には、研究者や生産者が土壌-植物-大気の連続体を測定するのを何十年も支援してきた経験があります。
葉面積指数は単一の数値であり、ある特定の時点における樹冠の統計的なスナップショットです。しかし、この一つの数値から重要な洞察が得られる可能性があります。
土壌水分の測定に必要な情報がすべてここに。
土壌の電気伝導率の測定について知っておく必要があることがすべてここにあります。

最新のコンテンツを定期的にお届けします。

